

Human Papillomavirus Type 16 Stimulates WAVE1- and WAVE2-Dependent Actin Protrusions for Endocytic Entry
- PMID: 40284985
- PMCID: PMC12031361
- DOI: 10.3390/v17040542
Human Papillomavirus Type 16 Stimulates WAVE1- and WAVE2-Dependent Actin Protrusions for Endocytic Entry
Abstract
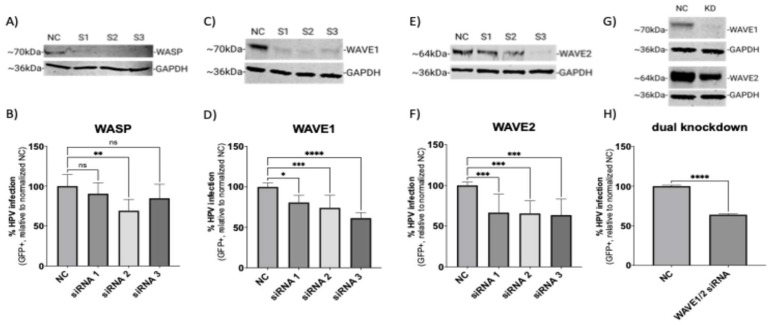
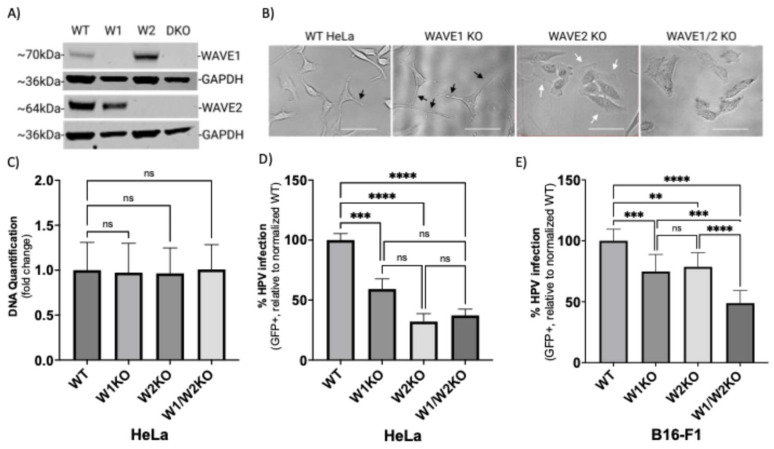
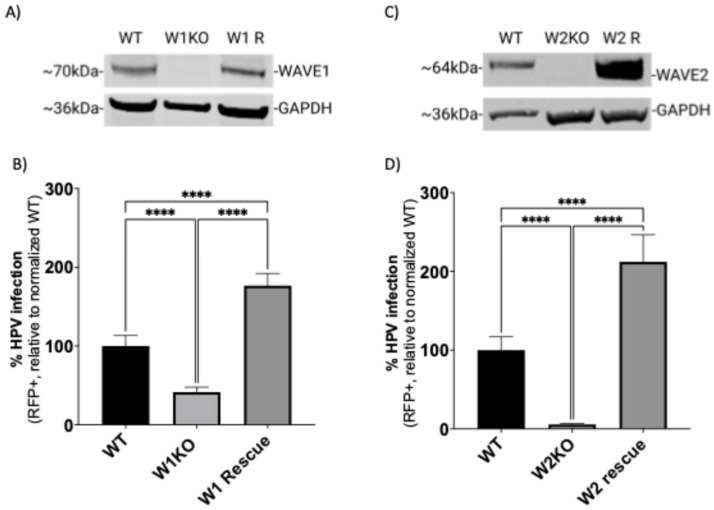
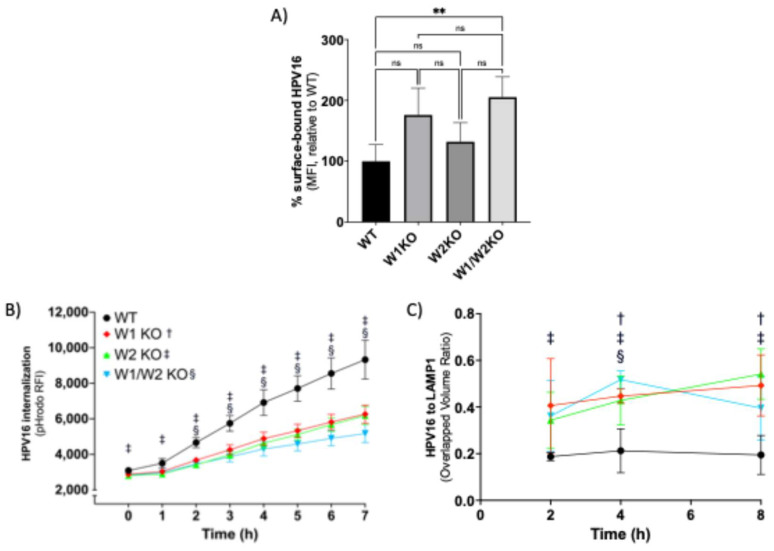
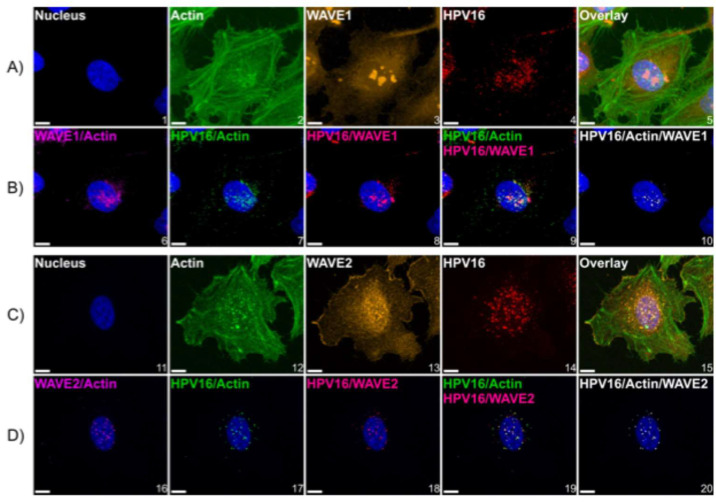
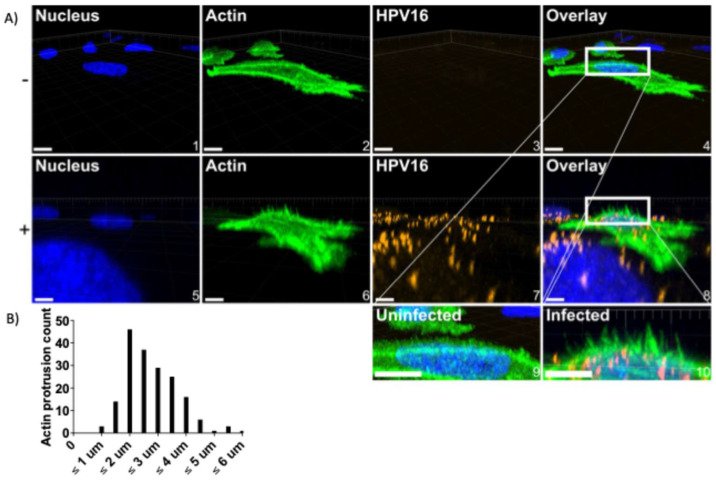
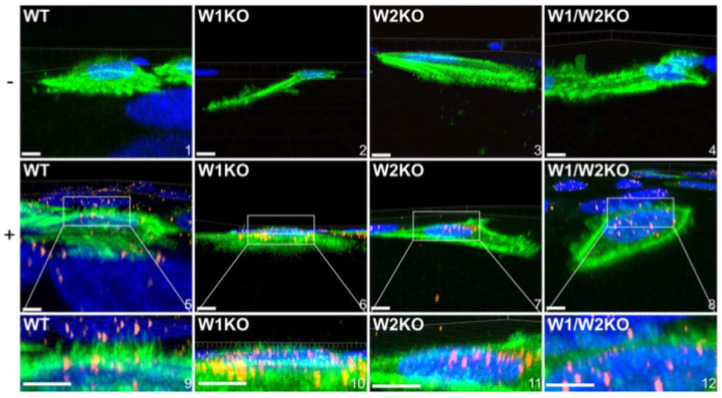
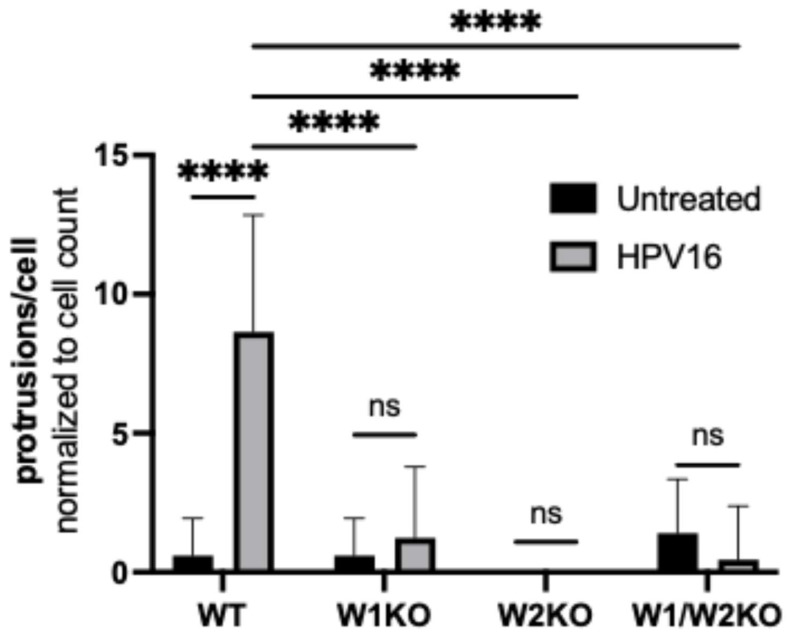
Human papillomavirus type 16 (HPV16) is an etiological agent of human cancers that requires endocytosis to initiate infection. HPV16 entry into epithelial cells occurs through a non-canonical endocytic pathway that is actin-driven, but it is not well understood how HPV16-cell surface interactions trigger actin reorganization in a way that facilitates entry. This study provides evidence that Wiskott-Aldrich syndrome protein family verprolin-homologous proteins 1 and 2 (WAVE1 and WAVE2) are molecular mediators of actin protrusions that occur at the cellular surface upon HPV addition to cells, and that this stimulation is a key step prior to endocytosis and intracellular trafficking. We demonstrate through post-transcriptional gene silencing and genome editing that WAVE1 and WAVE2 are critical for efficient HPV16 infection, and that restoration of each in knockout cells rescues HPV16 infection. Cells lacking WAVE1, WAVE2, or both internalize HPV16 at a significantly reduced rate. Microscopic analysis of fluorescently labeled cells revealed that HPV16, WAVE1, WAVE2, and actin are all colocalized at the cellular dorsal surface within a timeframe that precedes endocytosis. Within that same timeframe, we also found that HPV16-treated cells express cellular dorsal surface filopodia, which does not occur in cells lacking WAVE1 and WAVE2. Taken together, this study provides evidence that WAVE1 and WAVE2 mediate a key step prior to HPV entry into cells that involves actin reorganization in the form of cellular dorsal surface protrusions.
Keywords: HPV16; WAVE1; WAVE2; actin protrusions; human papillomavirus; viral entry.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Update of
-
WAVE1 and WAVE2 facilitate human papillomavirus-driven actin polymerization during cellular entry.bioRxiv [Preprint]. 2024 Oct 29:2024.10.28.620484. doi: 10.1101/2024.10.28.620484. bioRxiv. 2024. Update in: Viruses. 2025 Apr 08;17(4):542. doi: 10.3390/v17040542. PMID: 39553927 Free PMC article. Updated. Preprint.
References
-
- Bruni L., Albero G., Rowley J., Alemany L., Arbyn M., Giuliano A.R., Markowitz L.E., Broutet N., Taylor M. Global and regional estimates of genital human papillomavirus prevalence among men: A systematic review and meta-analysis. Lancet Glob. Health. 2023;11:e1345–e1362. doi: 10.1016/S2214-109X(23)00305-4. - DOI - PMC - PubMed
-
- Lang Kuhs K.A., Faden D.L., Chen L., Smith D.K., Pinheiro M., Wood C.B., Davis S., Yeager M., Boland J.F., Cullen M., et al. Genetic variation within the human papillomavirus type 16 genome is associated with oropharyngeal cancer prognosis. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2022;33:638–648. doi: 10.1016/j.annonc.2022.03.005. - DOI - PMC - PubMed
-
- Molano M., van den Brule A., Plummer M., Weiderpass E., Posso H., Arslan A., Meijer C.J.L.M., Muñoz N., Franceschi S., the HPV Study Group Determinants of Clearance of Human Papillomavirus Infections in Colombian Women with Normal Cytology: A Population-based, 5-Year Follow-up Study. Am. J. Epidemiol. 2003;158:486–494. doi: 10.1093/aje/kwg171. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
