

Metabolomic liquid biopsy dynamics predict early-stage HCC and actionable candidates of human hepatocarcinogenesis
- PMID: 40290517
- PMCID: PMC12023797
- DOI: 10.1016/j.jhepr.2025.101340
Metabolomic liquid biopsy dynamics predict early-stage HCC and actionable candidates of human hepatocarcinogenesis
Abstract
Background & aims: Actionable candidates of hepatocarcinogenesis remain elusive, and tools for early detection are suboptimal. Our aim was to demonstrate that serum metabolome profiles reflect the initiation of hepatocellular carcinoma (HCC) and enable the identification of biomarkers for early HCC detection and actionable candidates for chemoprevention.
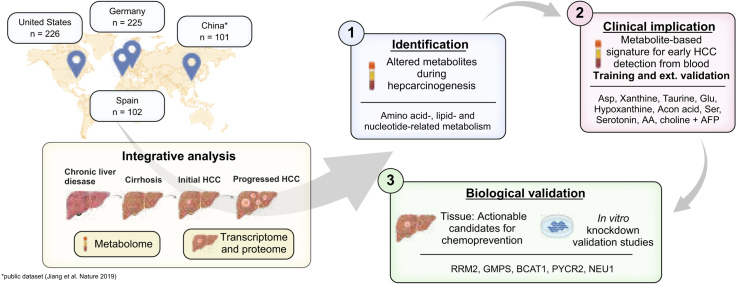
Methods: This global cohort study included 654 patients and 801 biospecimens. Following serum metabolome profiling across the spectrum of hepatocarcinogenesis, we conducted a phase II biomarker case-control study for early HCC detection. Findings were independently validated through in silico analysis, mRNA sequencing, and proteome profiling of primary HCC and non-tumoral tissue, and in vitro experiments.
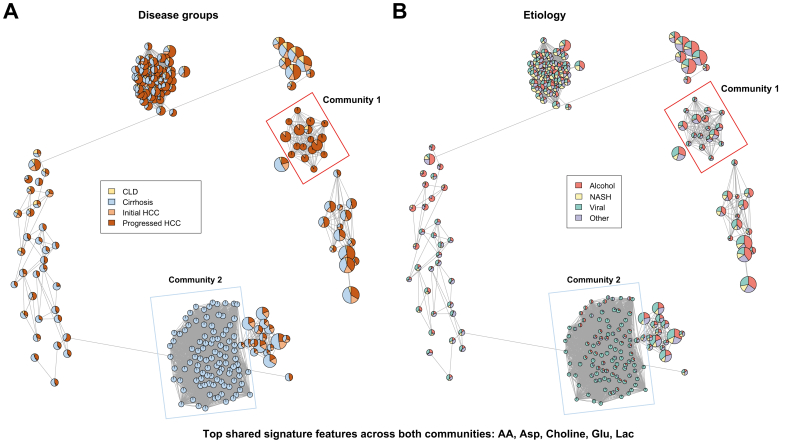
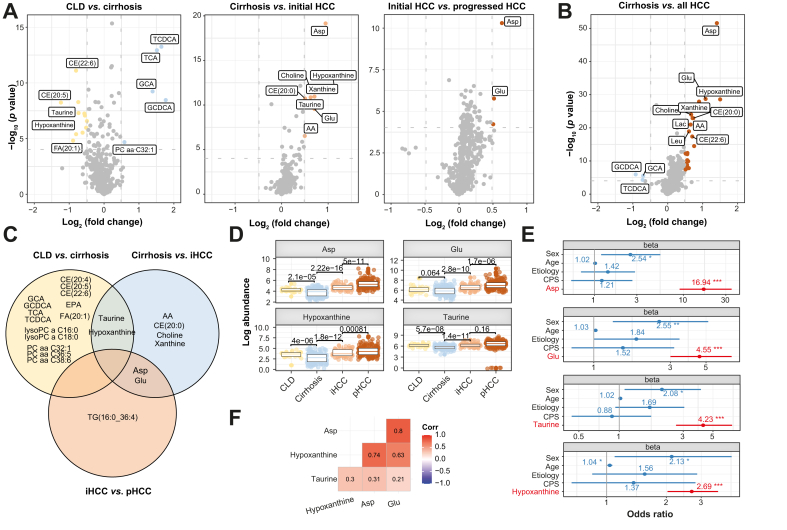
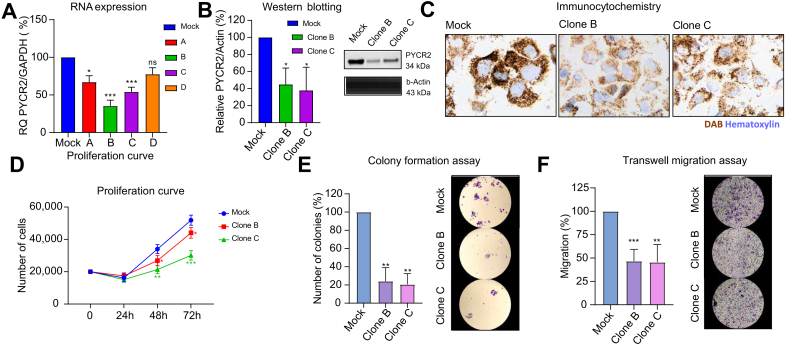
Results: Aspartic acid, glutamic acid, taurine, and hypoxanthine were differentially abundant in the serum across chronic liver disease, cirrhosis, initial HCC, and progressed HCC, independent of sex, age, and etiology. In a phase II biomarker case-control study, a blood-based metabolite signature yielded an AUC of 94% to discriminate between patients with early-stage HCC and controls with cirrhosis, including independent validation. Unsupervised biclustering (MoSBi), lipid network analysis (LINEX2), and pathway enrichment analysis confirmed alterations in amino acid-, lipid-, and nucleotide-related pathways. In tumor tissue, these pathways were significantly deregulated regarding gene and protein expression in two independent datasets, including actionable targets RRM2, GMPS, BCAT1, PYCR2, and NEU1. In vitro knockdown confirmed a functional role in proliferation and migration, as exemplified for PYCR2.
Conclusions: These findings demonstrate that serum metabolome profiling indicates deregulated metabolites and pathways during hepatocarcinogenesis. Our liquid biopsy approach accurately detects early-stage HCC outperforming currently recommended surveillance tools and facilitates identification of actionable candidates for chemoprevention.
Impact and implications: Deregulated cellular metabolism is a hallmark of cancer. In smaller studies, circulating metabolite profiles have been associated with HCC, although mainly in the context of fatty liver disease. Translation strategies for primary prevention or early detection are lacking. In this global study, we present an unsupervised landscape of the altered serum metabolome profile during hepatocarcinogenesis, independent of age, sex, and etiology. We provide a blood-based metabolite signature that accurately identifies early-stage HCC in a phase II biomarker study including independent validation. Further RRM2, GMPS, BCAT1, PYCR2, and NEU1 are identified in tumor tissue as actionable candidates for prevention. Our data provide the rationale for clinical trials testing liquid biopsy metabolome-based signatures for early HCC detection and the development of chemoprevention strategies.
Keywords: Early detection; Liver cancer; Metabolism; Prevention; Surveillance; Tumorigenesis.
© 2025 The Author(s).
Conflict of interest statement
KS has received advisory board fees from Roche, AstraZeneca, and MSD. CZ has received advisory board fees from Roche, MSD, and AstraZeneca. AW has received travel grants, honoraria, and/or advisory board fees from Bayer BMS, Sanofi, Roche, AstraZeneca, MSD, Merck KGaG, and Eisai. SH has received honoraria and/or consulting fees from Janssen Cilag, Ferring, AbbVie, Falk, Galapagos, Lilly, and BMS. BS has received financial support, fees, and/or grants from Astra Zeneca, BMS, Boston Scientific, Eisai, Incyte, MSD, Roche, Sanofi, and Sirtex Medical. JMB has received financial support, fees and/or grants from Albireo, Ipsen, Cymabay, AstraZeneca, Jazz Pharmaceuticals, Servier, Ikan Biotech, OWL Metabolomics, Incyte, Intercept, Advance, and Eisai. JUM has received grants, fees, and/or honoraria from AstraZeneca, MSD, Eisai, Ipsen, BMS, Incyte, and Roche. AV has received consulting fees from FirstWorld, Pioneering Medicine, and Genentech; and advisory board fees from BMS, Roche, Astra Zeneca, Eisai, and NGM Pharmaceuticals. He has stock options from Espervita and Atzeyo. JvF has received honoraria from Roche and AstraZeneca. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures





References
-
- Villanueva A. Hepatocellular carcinoma. N Engl J Med. 2019;380:1450–1462. - PubMed
-
- Llovet J.M., Kelley R.K., Villanueva A., et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021;7:6. - PubMed
-
- Craig A.J., von Felden J., Garcia-Lezana T., et al. Tumour evolution in hepatocellular carcinoma. Nat Rev Gastroenterol Hepatol. 2020;17:139–152. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
