

This is a preprint.
Identification of a stress-sensitive endogenous opioid-containing neuronal population in the paranigral ventral tegmental area
- PMID: 40291662
- PMCID: PMC12027071
- DOI: 10.1101/2025.04.08.647881
Identification of a stress-sensitive endogenous opioid-containing neuronal population in the paranigral ventral tegmental area
Abstract
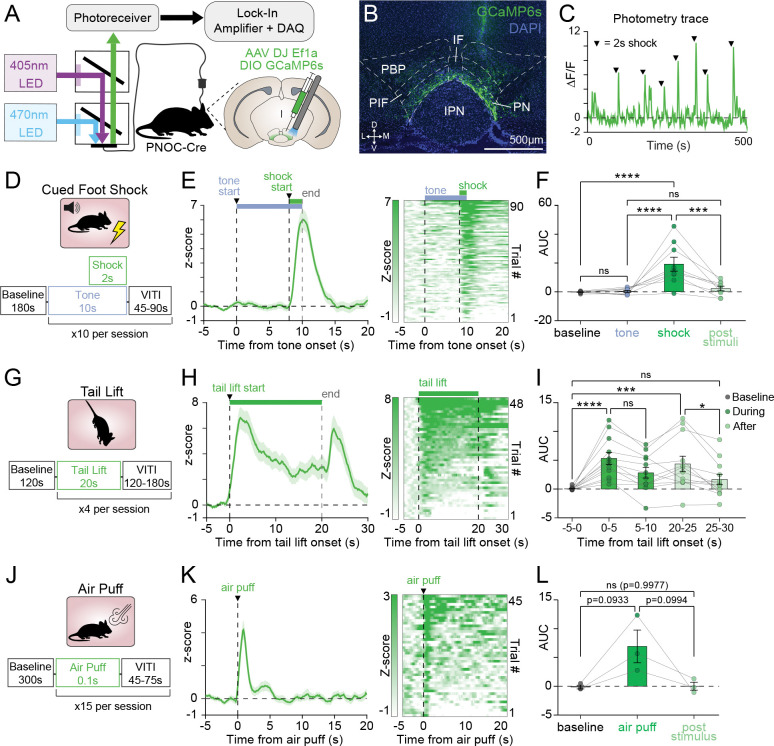
Nociceptin/orphanin FQ (N/OFQ), an endogenous opioid neuropeptide, and its G-protein coupled receptor NOPR have been implicated in motivation, feeding behaviors, and aversion. Stress-induced dysfunction in these states is central to the development of numerous psychiatric disorders, and the N/OFQ-NOPR system's role in reward- and stress-related responses has driven broad interest in NOPR as a therapeutic target for anxiety and depression. However, the impact of stress on N/OFQ signaling in the context of its influence on discrete midbrain reward circuitry remains unknown. To this end, we focused on a possible candidate population of N/OFQ neurons in the paranigral ventral tegmental area (pnVTAPNOC) that have been shown to act locally on NOPR-containing VTA dopamine neurons to suppress motivation. Here we report and characterize pnVTAPNOC sensitivity to stress exposure and identify a functional excitatory and inhibitory afferent input to this subpopulation from the lateral hypothalamus (LH). Our results indicate that pnVTAPNOC neurons become recruited during exposure to a range of acute stressor types, whereas the GABAergic input from the LH to this population is suppressed by predator odor stress, providing a mechanism for disinhibition of these neurons. These findings suggest that this N/OFQ population in the pnVTA could act as a critical bridge between stress and motivation through interactions with upstream hypothalamic circuitry.
Keywords: nociceptin; orphanin F/Q; stress.
Conflict of interest statement
COMPETING INTERESTS The authors declare no competing interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources