

This is a preprint.
Dynamic prefrontal coupling coordinates adaptive decision-making
- PMID: 40297698
- PMCID: PMC12036449
- DOI: 10.21203/rs.3.rs-6296852/v1
Dynamic prefrontal coupling coordinates adaptive decision-making
Abstract
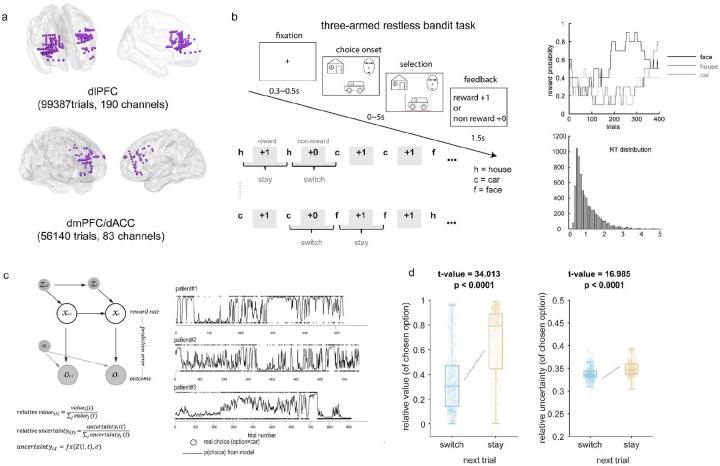
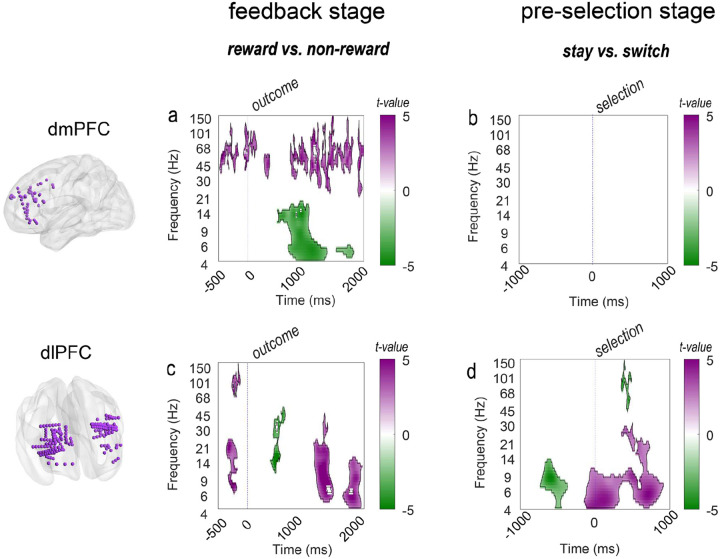
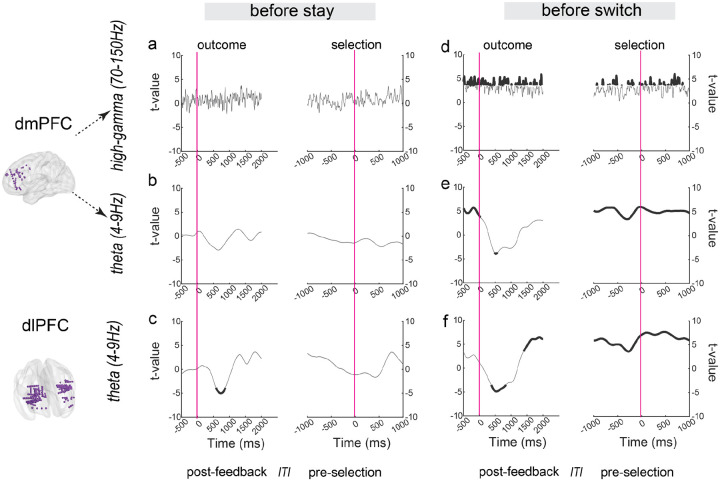
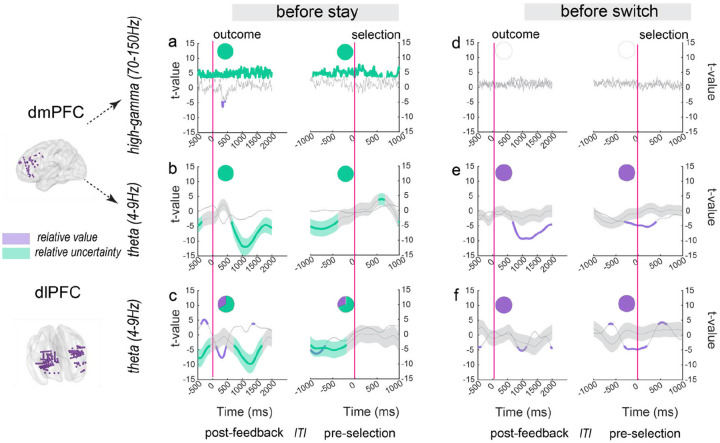
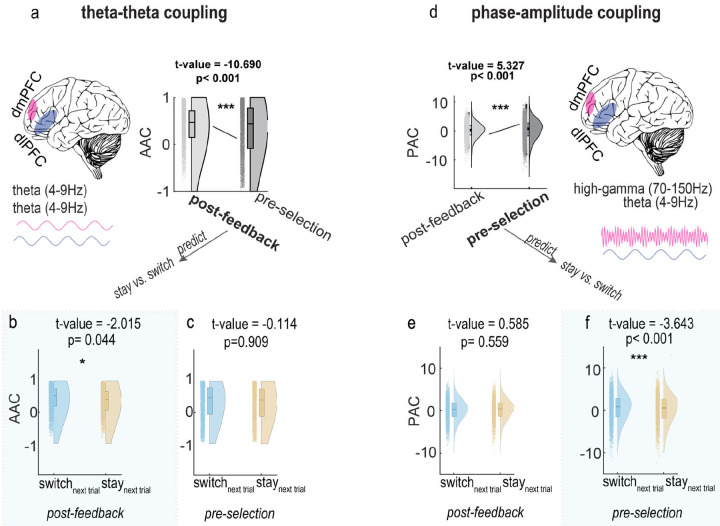
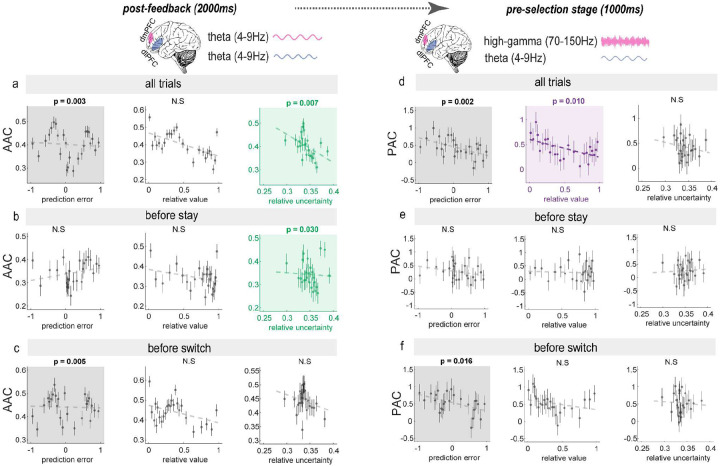
Adaptive decision-making requires flexibly maintaining or changing behavior in response to uncertainty. While the dorsomedial (dmPFC) and dorsolateral (dIPFC) prefrontal cortex are each essential for this ability, how they coordinate to drive adaptation remains unknown. Using intracranial EEG recordings from human participants performing a dynamic reward task, we identified distinct, frequency-specific computations: dmPFC high-gamma activity encoded uncertainty before stay decisions but transitioned to prediction error before switches, while theta activity shifted from uncertainty to value representation. In contrast, dIPFC theta activity signaled both value and uncertainty before stays, but predominantly value before switches. Crucially, these regions coordinated through two temporally specific coupling mechanisms that predicted behavioral changes: theta-theta amplitude coupling during feedback processing and theta-gamma phase coupling before decisions. Both coupling mechanisms strengthened before switches, suggesting that changing behavior requires greater dmPFC-dIPFC integration than maintaining. These findings reveal how the dorsal prefrontal cortex employs frequency-specific computations and precise temporal coordination to guide adaptive behavior.
Keywords: adaptation; dorsolateral prefrontal cortex; dorsomedial prefrontal cortex; inter-regional coupling; stay; switch; uncertainty; value.
Conflict of interest statement
Competing Interest Statement: The authors declare no competing interests.
Figures
Similar articles
-
Neural communication patterns underlying conflict detection, resolution, and adaptation.J Neurosci. 2014 Jul 30;34(31):10438-52. doi: 10.1523/JNEUROSCI.3099-13.2014. J Neurosci. 2014. PMID: 25080602 Free PMC article.
-
Neural dynamics of proactive and reactive cognitive control in medial and lateral prefrontal cortex.bioRxiv [Preprint]. 2025 Feb 13:2025.02.12.637987. doi: 10.1101/2025.02.12.637987. bioRxiv. 2025. PMID: 39990315 Free PMC article. Preprint.
-
Phase-Amplitude Coupling and Phase Synchronization Between Medial Temporal, Frontal and Posterior Brain Regions Support Episodic Autobiographical Memory Recall.Brain Topogr. 2022 Mar;35(2):191-206. doi: 10.1007/s10548-022-00890-4. Epub 2022 Jan 26. Brain Topogr. 2022. PMID: 35080692 Free PMC article.
-
Assessing the Effects of Continuous Theta Burst Stimulation Over the Dorsolateral Prefrontal Cortex on Human Cognition: A Systematic Review.Front Integr Neurosci. 2020 Aug 4;14:35. doi: 10.3389/fnint.2020.00035. eCollection 2020. Front Integr Neurosci. 2020. PMID: 32848648 Free PMC article.
-
Strategic control in decision-making under uncertainty.Eur J Neurosci. 2012 Apr;35(7):1075-82. doi: 10.1111/j.1460-9568.2012.08009.x. Eur J Neurosci. 2012. PMID: 22487037 Free PMC article. Review.
References
-
- Aberg K. C., Toren I. & Paz R. A neural and behavioral trade-off between value and uncertainty underlies exploratory decisions in normative anxiety. Mol. Psychiatry 27, 1573–1587 (March/2022). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
