

Effect of 1-DNJ on Oxidative Stress-Induced Apoptosis in Porcine Ovarian GCs Through Modulation of the PERK-ATF4/MFN2 Signaling Pathway
- PMID: 40298787
- PMCID: PMC12024285
- DOI: 10.3390/antiox14040456
Effect of 1-DNJ on Oxidative Stress-Induced Apoptosis in Porcine Ovarian GCs Through Modulation of the PERK-ATF4/MFN2 Signaling Pathway
Abstract
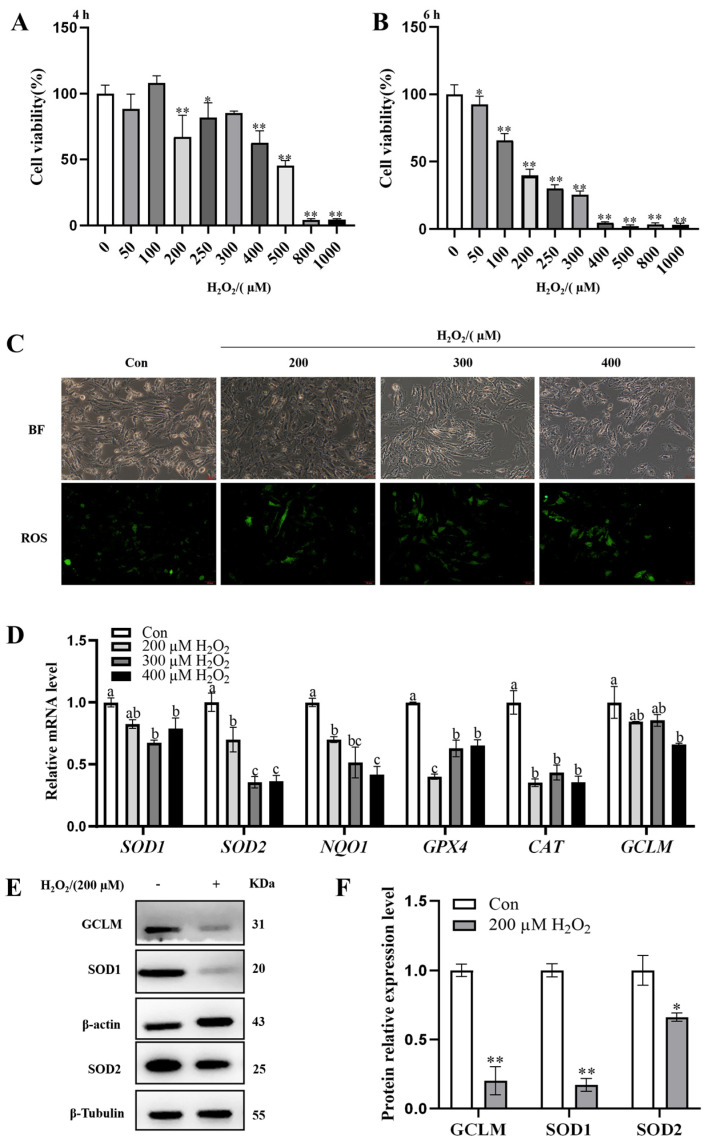
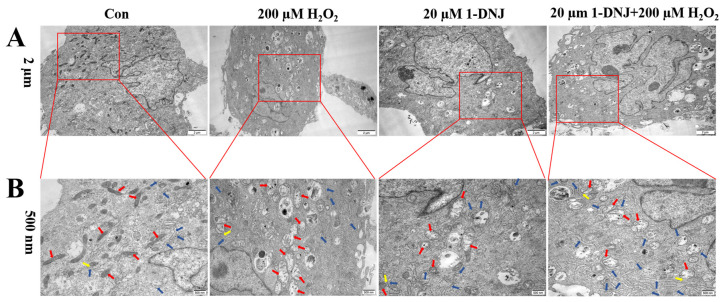
Oxidative stress (OS) is regarded as a major contributor to granulosa cellapoptosis in ovarian disease. 1-Deoxynojirimycin (1-DNJ), a naturally occurring plant alkaloid, exhibits antioxidant, anti-inflammatory, and metabolism-modulating properties. Mitochondria and endoplasmic reticulum (ER), crucial organelles regulating oxidative balance, interact through mitochondria-associated endoplasmic reticulum membranes (MAMs) for signaling and molecular exchange. However, it remains unclear whether 1-DNJ attenuates oxidative damage in ovarian granulosa cells (GCs) via MAMs-mediated ER-mitochondria crosstalk, which needs further exploration. This study aimed to investigate the mechanisms by which 1-DNJ affects oxidative damage and apoptosis induced by OS in porcine follicular GCs by regulating mitochondrial function, MAMs, and ER interactions. Here, we found that GCs suffered from OS, accompanied by the up-regulation of ROS and MDA, alongside reduced activity of antioxidant enzymes (CAT and T-SOD). Further studies revealed that the up-regulation of MAMs proteins (MFN2, MCU, and VDAC1) and pro-apoptosis proteins (BAX and Cleaved-capase3), along with increased mitochondrial ROS and Ca2+ levels, led to the down-regulation of MMP and ATP content. These, in turn, triggered mitochondrial dysfunction, and MAMs destabilization, and subsequent apoptosis. Additionally, the up-regulation of the protein levels of P-PERK/PERK, GRP78, ATF4, and CHOP protein expression activated the PERK-ATF4 signaling pathway, which triggered endoplasmic reticulum stress (ERS). Conversely, 1-DNJ alleviated H2O2-induced mitochondrial and MAMs dysfunction and ERS, which in turn attenuated apoptosis. Further, ATF4 knockdown inhibited MFN2 protein expression, which attenuated H2O2-induced MMP inhibition, Ca2+ overload, ROS production, and mitochondrial damage. In summary, 1-DNJ mitigated OS-induced mitochondrial dysfunction in GCs and regulated ER-mitochondrial communication through MAMs, reducing OS-induced apoptosis. The present study demonstrates that 1-DNJ protects ovarian GCs from OS-induced damage by modulating ER and mitochondrial homeostasis through MAMs, offering new perspectives and a theoretical basis for the treatment of ovarian diseases.
Keywords: 1-DNJ; endoplasmic reticulum stress; mitochondria-associated endoplasmic reticulum membrane; mitochondrial dysfunction; ovarian granulosa cells; oxidative stress.
Conflict of interest statement
No conflicts of interest exist in the submission of this manuscript, and the manuscript is approved by all authors for publication. I would like to declare on behalf of my coauthors that the work described was original research that has not been published previously, and is not under consideration for publication elsewhere, in whole or in part. All of the authors listed have approved the manuscript that is enclosed.
Figures










References
-
- Duan H., Yang S., Yang S., Zeng J., Yan Z., Zhang L., Ma X., Dong W., Zhang Y., Zhao X., et al. The mechanism of curcumin to protect mouse ovaries from oxidative damage by regulating AMPK/mTOR mediated autophagy. Phytomedicine. 2024;128:155468. - PubMed
-
- Wang Y., Pattarawat P., Zhang J., Kim E., Zhang D., Fang M., Jannaman E.A., Yuan Y., Chatterjee S., Kim J.J., et al. Effects of Cyanobacterial Harmful Algal Bloom Toxin Microcystin-LR on Gonadotropin-Dependent Ovarian Follicle Maturation and Ovulation in Mice. Environ. Health Perspect. 2023;131:67010. - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
