

Bikaverin as a molecular weapon: enhancing Fusarium oxysporum pathogenicity in bananas via rhizosphere microbiome manipulation
- PMID: 40301992
- PMCID: PMC12042607
- DOI: 10.1186/s40168-025-02109-7
Bikaverin as a molecular weapon: enhancing Fusarium oxysporum pathogenicity in bananas via rhizosphere microbiome manipulation
Abstract
Background: Fusarium wilt, caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4), poses a severe threat to global banana production. Secondary metabolites are critical tools employed by pathogens to interact with their environment and modulate host-pathogen dynamics. Bikaverin, a red-colored polyketide pigment produced by several Fusarium species, has been studied for its pharmacological properties, but its ecological roles and impact on pathogenicity remain unclear.
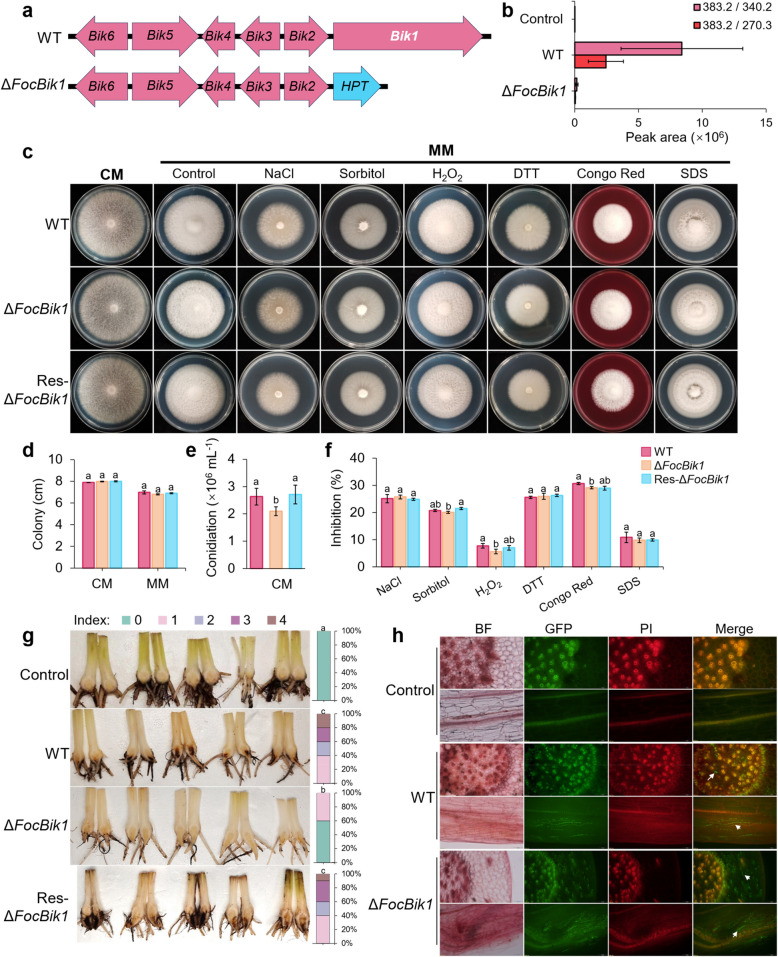
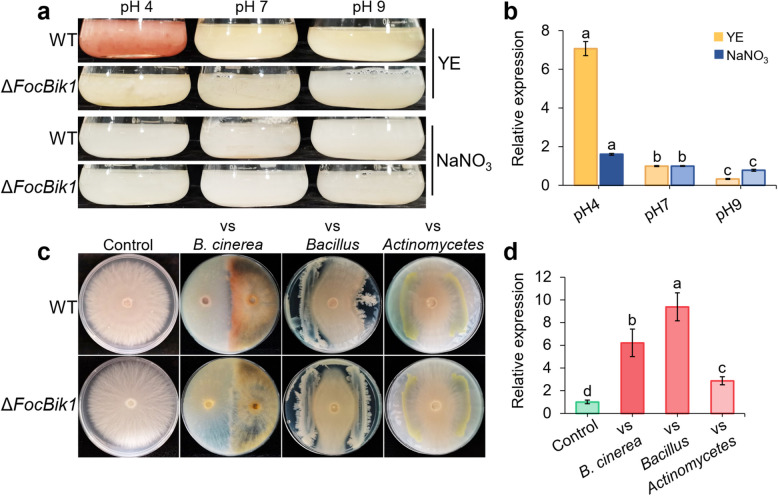
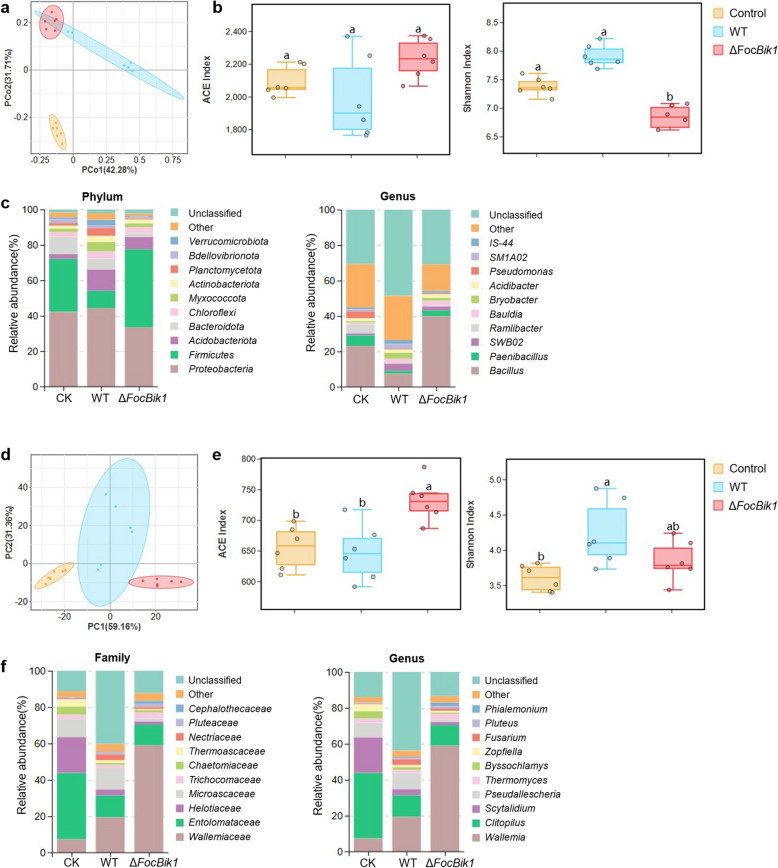
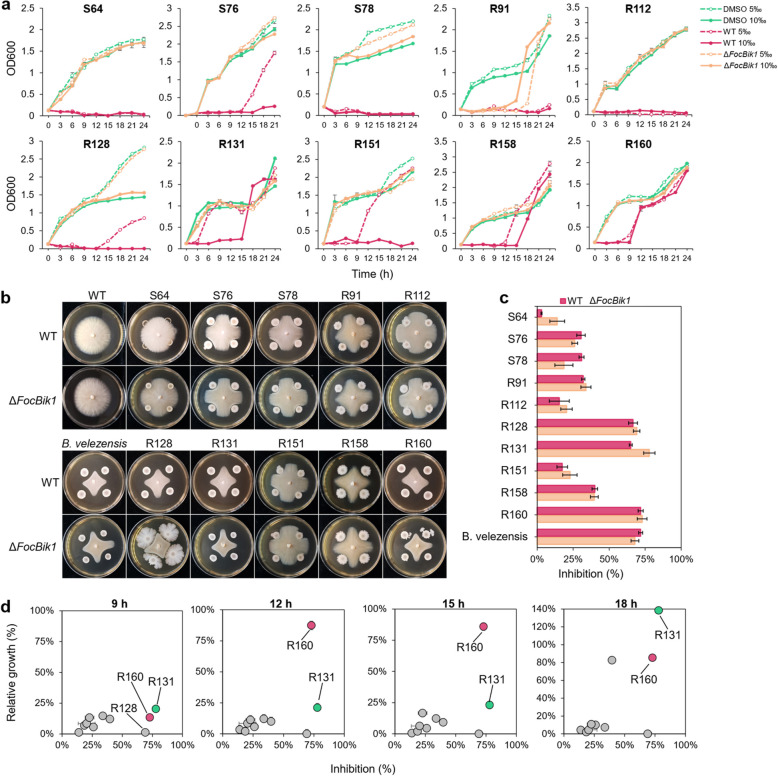
Results: This study investigated the role of bikaverin in Foc TR4, focusing on its contribution to pathogenicity and its interaction with the rhizosphere microbiome. Pathogenicity assays under sterile and autoclaved conditions demonstrated that bikaverin does not directly contribute to pathogenicity by affecting the infection process or damaging host tissues. Instead, bikaverin indirectly enhances Foc TR4's pathogenicity by reshaping the rhizosphere microbiome. It suppresses beneficial plant growth-promoting rhizobacteria, such as Bacillus, while promoting the dominance of fungal genera, thereby creating a microbial environment beneficial for pathogen colonization and infection. Notably, bikaverin biosynthesis was found to be tightly regulated by environmental cues, including acidic pH, nitrogen scarcity, and microbial competition. Co-culture with microbes such as Bacillus velezensis and Botrytis cinerea strongly induced bikaverin production and upregulated expression of the key bikaverin biosynthetic gene FocBik1. In addition, the identification of bikaverin-resistant Bacillus BR160, a strain with broad-spectrum antifungal activity, highlights its potential as a biocontrol agent for banana wilt management, although its stability and efficiency under field conditions require further validation.
Conclusions: Bikaverin plays an indirect yet important role in the pathogenicity of Foc TR4 by manipulating the rhizosphere microbiome. This ecological function underscores its potential as a target for sustainable disease management strategies. Future research should focus on elucidating the molecular mechanisms underlying bikaverin-mediated microbial interactions, using integrated approaches such as transcriptomics and metabolomics. Together, these findings provide a foundation for novel approaches to combat banana wilt disease and enhance crop resistance. Video Abstract.
Keywords: Bacillus; Banana wilt disease; Bikaverin; Foc TR4; Plant growth-promoting rhizobacteria; Rhizosphere microbiome; Secondary metabolites.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures
References
-
- Ploetz RC. Panama disease: an old nemesis rears its ugly head: part 1. The beginnings of the banana export trades. Plant Health Prog. 2005;6(1):18.
-
- Ismaila AA, Ahmad K, Siddique Y, Wahab MAA, Kutawa AB, Abdullahi A, et al. Fusarium wilt of banana: current update and sustainable disease control using classical and essential oils approaches. Hortic Plant J. 2023;9(1):1–28. - DOI
-
- Fu L, Penton CR, Ruan Y, Shen Z, Xue C, Li R, Shen Q. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol Biochem. 2017;104:39–48. - DOI
MeSH terms
Substances
Supplementary concepts
Grants and funding
- ZDYF2024XDNY214/Key Research and Development Project of Hainan Province
- 32001846, 32260042, 32160594/National Natural Science Foundation of China
- 32001846, 32260042, 32160594/National Natural Science Foundation of China
- SCKJ-JYRC-2024-15/Project of Sanya Yazhou Bay Science and Technology City, China
LinkOut - more resources
Full Text Sources
