

Cgref1 is a CREB-H-regulated hepatokine that promotes hepatic de novo lipogenesis by mediating epididymal fat insulin resistance
- PMID: 40303310
- PMCID: PMC12035884
- DOI: 10.7150/ijbs.97008
Cgref1 is a CREB-H-regulated hepatokine that promotes hepatic de novo lipogenesis by mediating epididymal fat insulin resistance
Abstract
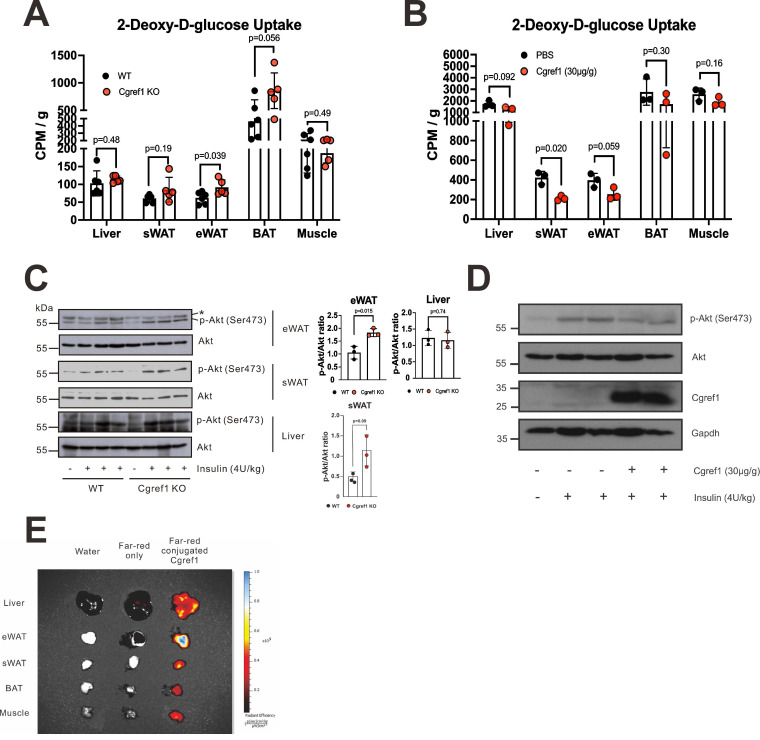
Rationale: Type 2 diabetes mellitus and metabolic dysfunction-associated steatotic liver disease (MASLD) are interrelated metabolic disorders that pose significant health concerns. Hepatokines and other regulatory factors implicated in these diseases are incompletely understood. Here, we report on a new hepatokine named cell growth regulator with EF-hand domain 1 (Cgref1) that modulates lipid metabolism to aggravate these conditions. Methods: Cgref1 was identified by microarray analysis of downregulated genes in liver of Creb3l3 -/- mice. Cgref1 -/- mice were subjected to transcriptomic, metabolomic and lipidomic analyses as well as metabolic assays. Gain-of-function and loss-of-function assays were performed in primary hepatocytes and cultured human and mouse cells. Results: Cgref1 expression is induced by hepatic transcription factor CREB-H. Secreted Cgref1 primarily targets epididymal white adipose tissue (eWAT), where insulin signalling and glucose uptake are suppressed. Cgref1-/- mice showed lower tendencies of developing obesity, hyperglycaemia and dyslipidaemia, associated with compromised hepatic de novo lipogenesis. Thus, Cgref1 poses an advantage to maintain the normal functioning of vital organs by preserving glucose from being absorbed into eWAT. However, in circumstances where Cgref1 expression becomes excessive, eWAT develops insulin resistance, which in turn promotes hepatic glucose production, lipogenesis and MASLD development. Conclusion: As a hepatokine that affects blood glucose levels and lipogenesis, Cgref1 is a potential target in the intervention of metabolic disorders.
Keywords: CREB-H transcription factor; Cgref1; diabetes; hepatokine; metabolic dysfunction-associated steatotic liver disease (MASLD); metabolic syndrome.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures






Similar articles
-
The adipokine sFRP4 induces insulin resistance and lipogenesis in the liver.Biochim Biophys Acta Mol Basis Dis. 2019 Oct 1;1865(10):2671-2684. doi: 10.1016/j.bbadis.2019.07.008. Epub 2019 Jul 20. Biochim Biophys Acta Mol Basis Dis. 2019. PMID: 31336149
-
Hepatic CREBZF couples insulin to lipogenesis by inhibiting insig activity and contributes to hepatic steatosis in diet-induced insulin-resistant mice.Hepatology. 2018 Oct;68(4):1361-1375. doi: 10.1002/hep.29926. Epub 2018 May 18. Hepatology. 2018. PMID: 29637572
-
Interleukin-1α deficiency reduces adiposity, glucose intolerance and hepatic de-novo lipogenesis in diet-induced obese mice.BMJ Open Diabetes Res Care. 2019 Oct 17;7(1):e000650. doi: 10.1136/bmjdrc-2019-000650. eCollection 2019. BMJ Open Diabetes Res Care. 2019. PMID: 31749969 Free PMC article.
-
Adipose tissue regulates insulin sensitivity: role of adipogenesis, de novo lipogenesis and novel lipids.J Intern Med. 2016 Nov;280(5):465-475. doi: 10.1111/joim.12540. Epub 2016 Oct 3. J Intern Med. 2016. PMID: 27699898 Free PMC article. Review.
-
Hepatic steatosis: a role for de novo lipogenesis and the transcription factor SREBP-1c.Diabetes Obes Metab. 2010 Oct;12 Suppl 2:83-92. doi: 10.1111/j.1463-1326.2010.01275.x. Diabetes Obes Metab. 2010. PMID: 21029304 Review.
References
-
- Younossi ZM, Golabi P, de Avila L, Paik JM, Srishord M, Fukui N. et al. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J Hepatol. 2019;71(4):793–801. - PubMed
-
- Utzschneider KM, Kahn SE. Review: The role of insulin resistance in nonalcoholic fatty liver disease. J Clin Endocrinol Metab. 2006;91(12):4753–61. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
