

PRMT5 promotes full-length HTT expression by repressing multiple proximal intronic polyadenylation sites
- PMID: 40304179
- PMCID: PMC12041856
- DOI: 10.1093/nar/gkaf347
PRMT5 promotes full-length HTT expression by repressing multiple proximal intronic polyadenylation sites
Abstract
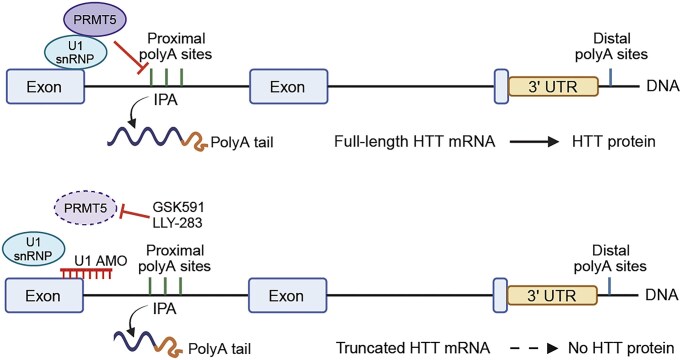
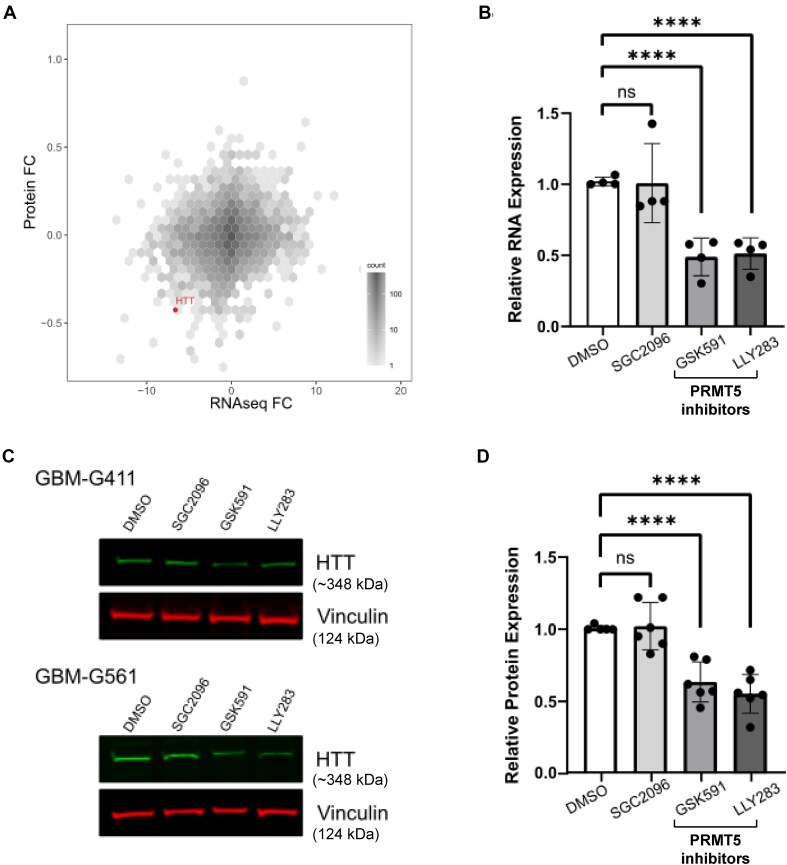
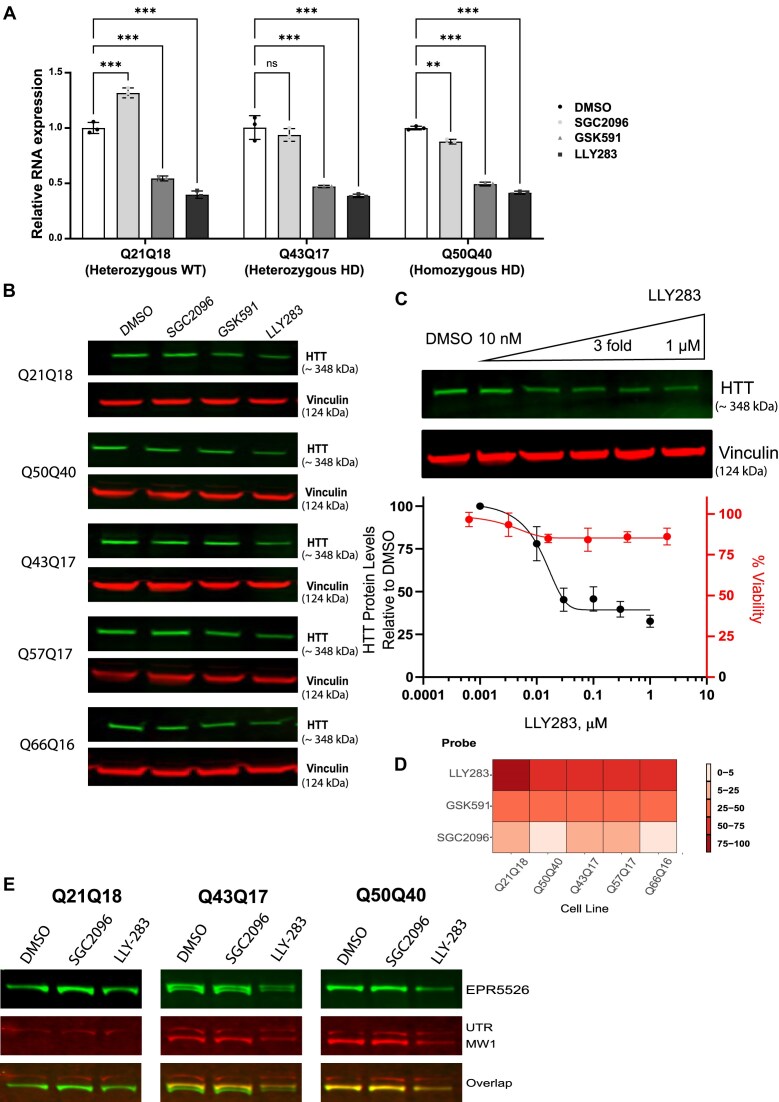
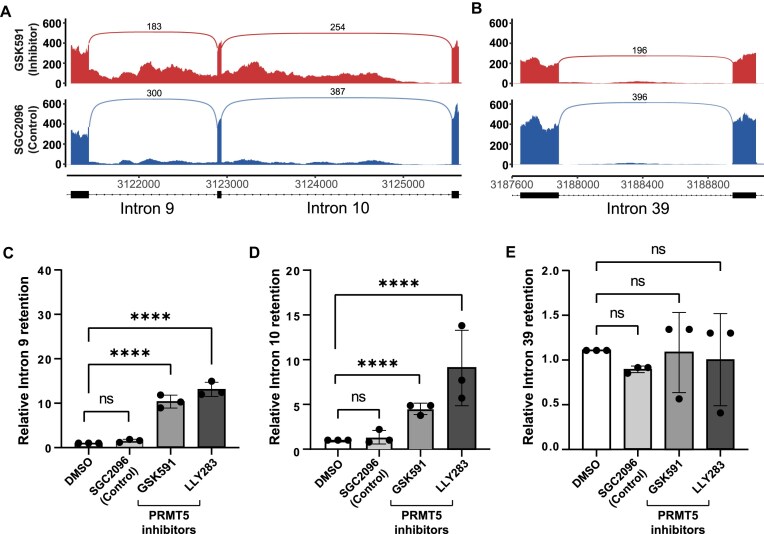
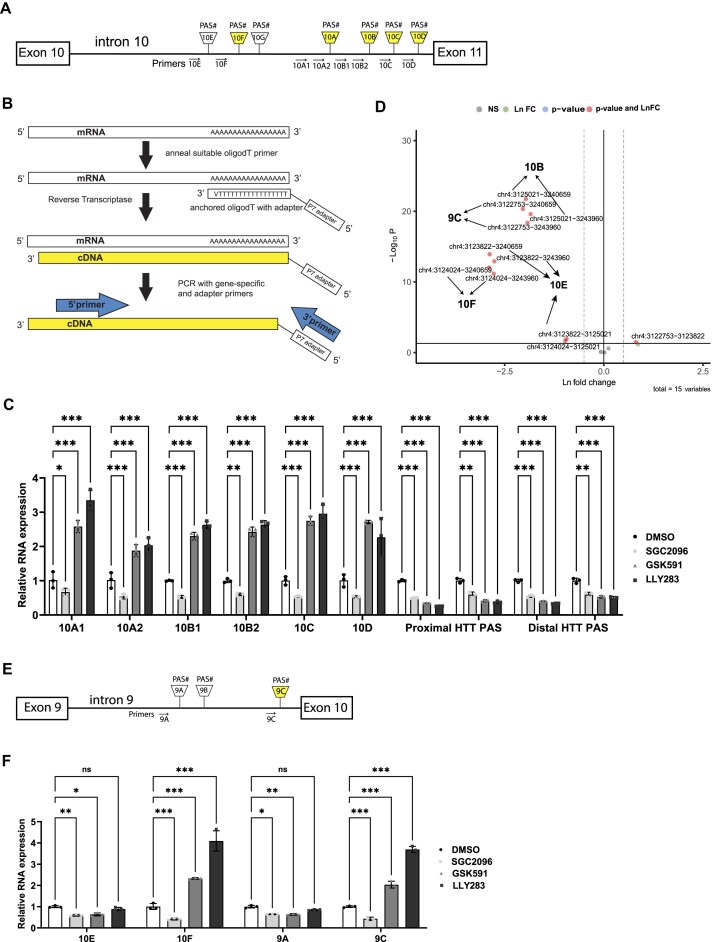
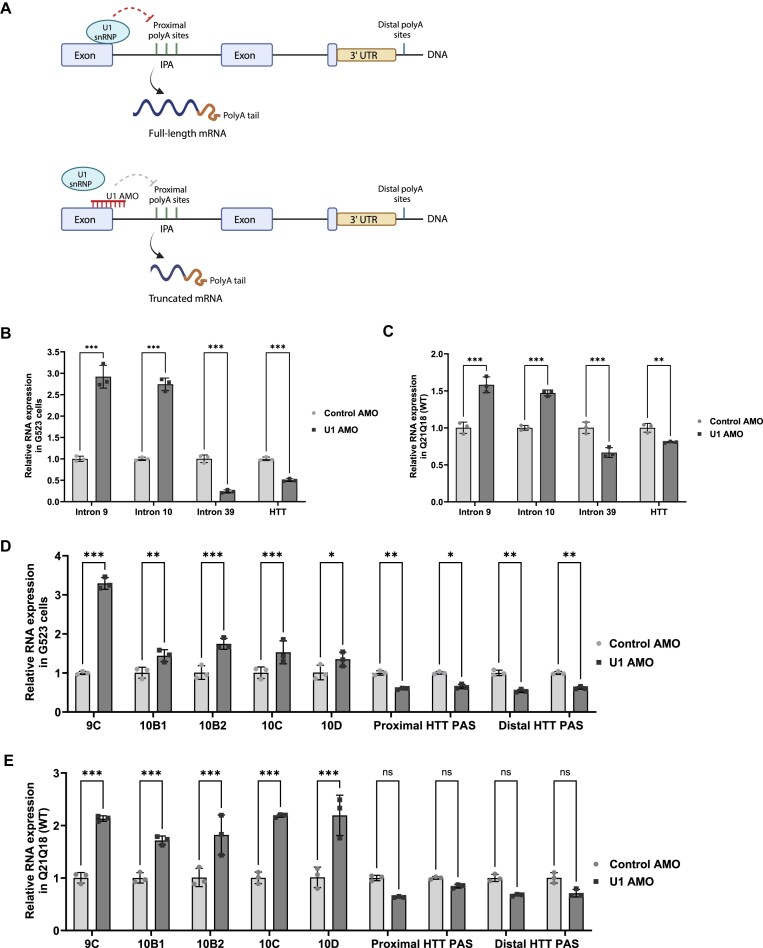
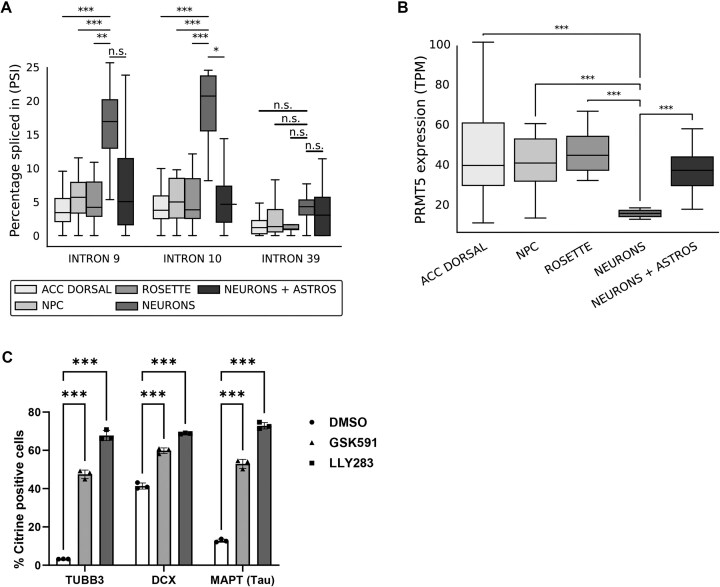
Expansion of the CAG trinucleotide repeat tract in exon 1 of the Huntingtin (HTT) gene causes Huntington's disease (HD) through the expression of a polyglutamine-expanded form of the HTT protein. This mutation triggers cellular and biochemical pathologies, leading to cognitive, motor, and psychiatric symptoms in HD patients. Targeting HTT splicing with small molecule drugs is a compelling approach to lowering HTT protein levels to treat HD, and splice modulators are currently being tested in the clinic. Here, we identify PRMT5 as a novel regulator of HTT messenger RNA (mRNA) splicing and alternative polyadenylation. PRMT5 inhibition disrupts the splicing of HTT introns 9 and 10, leading to the activation of multiple proximal intronic polyadenylation sites within these introns and promoting premature termination, cleavage, and polyadenylation of the HTT mRNA. This suggests that HTT protein levels may be lowered due to this mechanism. We also detected increasing levels of these truncated HTT transcripts across a series of neuronal differentiation samples, which correlated with lower PRMT5 expression. Notably, PRMT5 inhibition in glioblastoma stem cells potently induced neuronal differentiation. We posit that PRMT5-mediated regulation of intronic polyadenylation, premature termination, and cleavage of the HTT mRNA modulates HTT expression and plays an important role during neuronal differentiation.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
None declared.
Figures
References
MeSH terms
Substances
Grants and funding
- Boehringer Ingelheim
- SU2C-AACR-DT-19-15/Stand Up to Cancer Canada
- OGI-196/Ontario Genomics Institute
- Bayer AG
- Genome Canada
- Structural Genomics Consortium
- Ontario Institute for Cancer Research
- Princess Margaret Postdoctoral Fellowship
- Merck KGaA
- Genentech
- Irma T. Hirschl Trust
- R01 GM108646/GM/NIGMS NIH HHS/United States
- 142434/Canadian Institute of Health Research
- American Association for Cancer Research
- MITACS
- Bristol Myers Squibb
- Takeda
- R01GM108646/GF/NIH HHS/United States
- Janssen
- Government of Ontario
- Pfizer
- Entertainment Industry Foundation
LinkOut - more resources
Full Text Sources
