

Stress-dependent activation of the Listeria monocytogenes virulence program ensures bacterial resilience during infection
- PMID: 40304513
- PMCID: PMC12153296
- DOI: 10.1128/mbio.00719-25
Stress-dependent activation of the Listeria monocytogenes virulence program ensures bacterial resilience during infection
Abstract
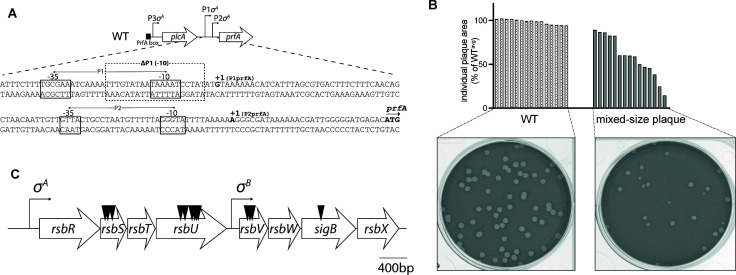
Listeria monocytogenes (Lm) is a Gram-positive, facultative intracellular pathogen that uses both a housekeeping (P1) and stress-activated (Sigma B-dependent) promoter (P2) to express the master virulence regulator PrfA. The Sigma B regulon contains over 300 genes known to respond to different stressors. However, the role of Sigma B in the regulation of prfA during the infection remains uncertain. To define pathways that lead to Sigma B-dependent prfA activation, we performed a genetic screen in L2 fibroblasts using ΔP1 Lm that only has the Sigma B-dependent promoter directly upstream of prfA. The screen identified transposon insertions in a large bacterial sensory organelle known as the stressosome. The absence of functional stressosome components resulted in heterogeneity within bacterial populations, with some bacteria behaving like wild type, while other members of the population exhibited defects in either vacuolar escape and/or cell-to-cell spread. We show that the heterogeneity of the stressosome mutants cannot be rescued by constitutive activation of PrfA. These data defined the importance of the stressosome in controlling bacterial homogeneity and characterized the function of the stressosome in robust virulence activation during infection. ΔP1 Lm model provides new opportunities to identify host-specific signals necessary for stressosome-dependent signaling during Listeria pathogenesis.IMPORTANCEMicrobial pathogens must adapt to varying levels of stress to survive. This study uncovered a link between stress sensing and activation of the virulence program in a facultative intracellular pathogen, Listeria monocytogenes. We show that host-imposed stress is sensed by the signaling machinery known as the stressosome to ensure robust and resilient virulence responses in vivo. Stressosome-dependent activation of the master virulence regulator PrfA was necessary to maintain L. monocytogenes homogeneity within the bacteria population during the transition between early and late stages of intracellular infection. This work also provides a model to further characterize how specific stress stimuli affect bacterial survival within the host, which is critical for our understanding of bacterial pathogenesis.
Keywords: intracellular bacteria; macrophages; pathogenesis; sigma factors; stress adaptation; virulence regulation.
Conflict of interest statement
D.A.P. has a financial interest in Laguna Biotherapeutics, and both he and the company could benefit from the commercialization of the results of this research.
Figures





Similar articles
-
Contributions of Listeria monocytogenes sigmaB and PrfA to expression of virulence and stress response genes during extra- and intracellular growth.Microbiology (Reading). 2006 Jun;152(Pt 6):1827-1838. doi: 10.1099/mic.0.28758-0. Microbiology (Reading). 2006. PMID: 16735745
-
Flick of a switch: regulatory mechanisms allowing Listeria monocytogenes to transition from a saprophyte to a killer.Microbiology (Reading). 2019 Aug;165(8):819-833. doi: 10.1099/mic.0.000808. Epub 2019 May 20. Microbiology (Reading). 2019. PMID: 31107205 Review.
-
Cross Talk between SigB and PrfA in Listeria monocytogenes Facilitates Transitions between Extra- and Intracellular Environments.Microbiol Mol Biol Rev. 2019 Sep 4;83(4):e00034-19. doi: 10.1128/MMBR.00034-19. Print 2019 Nov 20. Microbiol Mol Biol Rev. 2019. PMID: 31484692 Free PMC article. Review.
-
Transcriptomic and phenotypic analyses identify coregulated, overlapping regulons among PrfA, CtsR, HrcA, and the alternative sigma factors sigmaB, sigmaC, sigmaH, and sigmaL in Listeria monocytogenes.Appl Environ Microbiol. 2011 Jan;77(1):187-200. doi: 10.1128/AEM.00952-10. Epub 2010 Oct 29. Appl Environ Microbiol. 2011. PMID: 21037293 Free PMC article.
-
Sigma B contributes to PrfA-mediated virulence in Listeria monocytogenes.Infect Immun. 2002 Jul;70(7):3948-52. doi: 10.1128/IAI.70.7.3948-3952.2002. Infect Immun. 2002. PMID: 12065541 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
