

Inclusion of a retroviral protease enhances the immunogenicity of VLP-forming mRNA vaccines against HIV-1 or SARS-CoV-2 in mice
- PMID: 40305570
- PMCID: PMC12151460
- DOI: 10.1126/scitranslmed.adt9576
Inclusion of a retroviral protease enhances the immunogenicity of VLP-forming mRNA vaccines against HIV-1 or SARS-CoV-2 in mice
Abstract
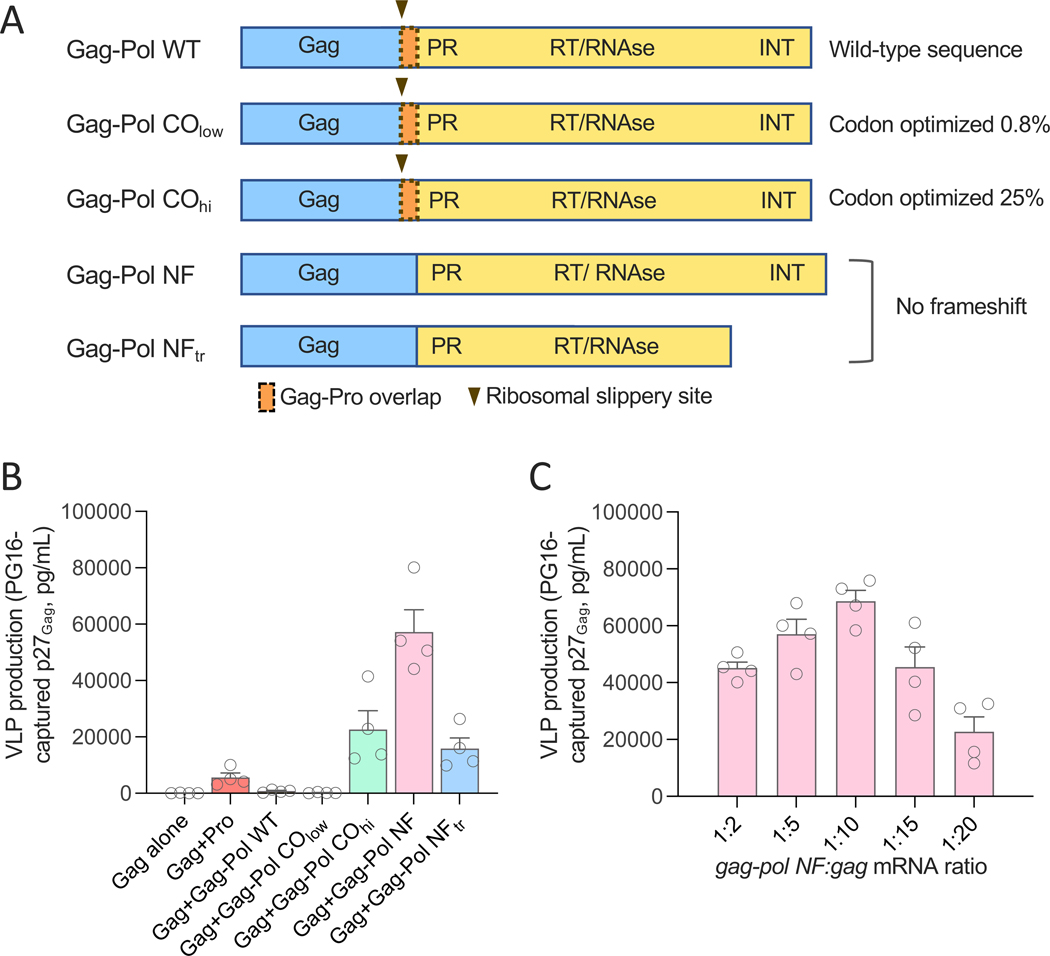
Messenger RNA (mRNA) has emerged as a highly effective and versatile platform for vaccine delivery. We previously designed a virus-like particle (VLP)-forming env-gag mRNA vaccine against human immunodeficiency virus-1 (HIV-1) that elicited envelope-specific neutralizing antibodies and protection from heterologous simian-human immunodeficiency virus (SHIV) infection in rhesus macaques. Here, we introduce a key technological advance to this platform by inclusion of mRNA encoding a retroviral protease to process Gag and produce mature VLPs. Appropriately dosed and timed expression of the protease was achieved using a full-length gag-pol mRNA transcript. Addition of gag-pol mRNA to an HIV-1 env-gag mRNA vaccine resulted in enhanced titers of envelope trimer-binding and neutralizing antibodies in a mouse model. Analogous results were obtained with a hybrid Gag-based, VLP-forming severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) mRNA vaccine expressing an engineered spike protein. Thus, inclusion of a retroviral protease can increase the immunogenicity of Gag-based, VLP-forming mRNA vaccines against human pathogens.
Conflict of interest statement
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
