

Altered extracellular matrix structure and elevated stiffness in a brain organoid model for disease
- PMID: 40312467
- PMCID: PMC12045990
- DOI: 10.1038/s41467-025-59252-w
Altered extracellular matrix structure and elevated stiffness in a brain organoid model for disease
Abstract
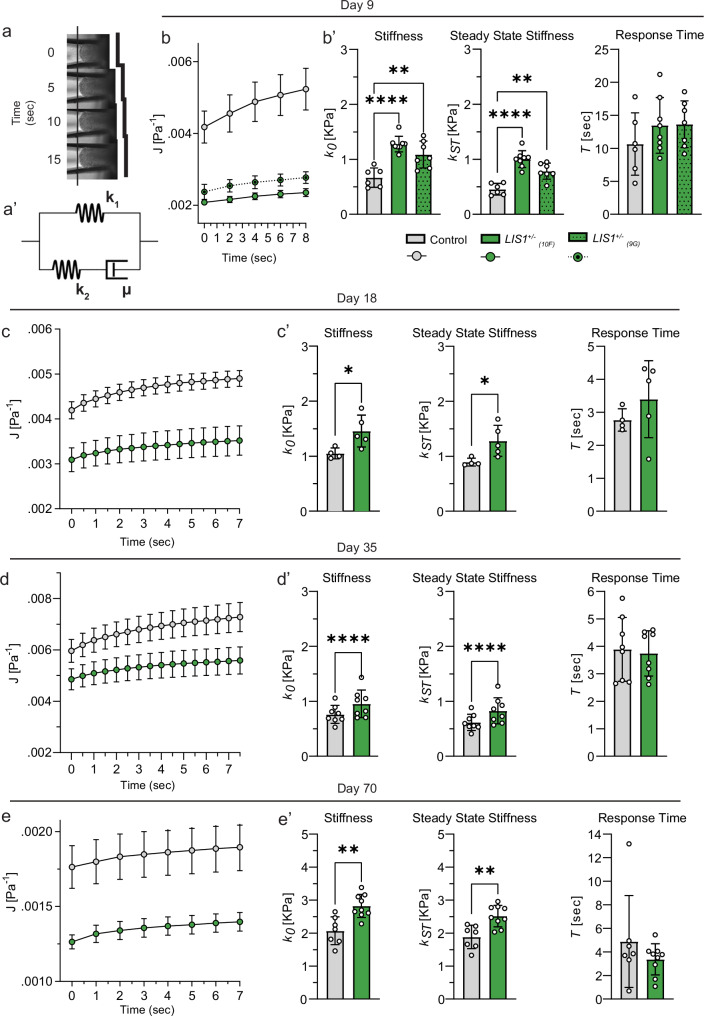
The viscoelastic properties of tissues influence their morphology and cellular behavior, yet little is known about changes in these properties during brain malformations. Lissencephaly, a severe cortical malformation caused by LIS1 mutations, results in a smooth cortex. Here, we show that human-derived brain organoids with LIS1 mutation exhibit increased stiffness compared to controls at multiple developmental stages. This stiffening correlates with abnormal extracellular matrix (ECM) expression and organization, as well as elevated water content, measured by diffusion-weighted MRI. Short-term MMP9 treatment reduces both stiffness and water diffusion levels to control values. Additionally, a computational microstructure mechanical model predicts mechanical changes based on ECM organization. These findings suggest that LIS1 plays a critical role in ECM regulation during brain development and that its mutation leads to significant viscoelastic alterations.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




References
-
- Franze, K. The mechanical control of nervous system development. Development140, 3069–3077 (2013). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
