

Paradoxical Activation of Entheseal Myeloid Cells by JAK1 and Tyk2 Inhibitors via Interleukin-10 Antagonism
- PMID: 40320905
- PMCID: PMC12479184
- DOI: 10.1002/art.43210
Paradoxical Activation of Entheseal Myeloid Cells by JAK1 and Tyk2 Inhibitors via Interleukin-10 Antagonism
Abstract
Objective: JAK inhibition (JAKi) is effective in seronegative spondyloarthropathy (SpA) spectrum disorders, but Tyk2 inhibition failed in SpA spectrum ulcerative colitis, and tofacitinib showed minimal benefit in Crohn disease, which highlights the complex role for JAK/STAT signaling in different inflammatory processes. In this study, we investigated whether JAKi might paradoxically activate entheseal innate immunity and aimed to identify the key regulatory cytokines involved in this process.
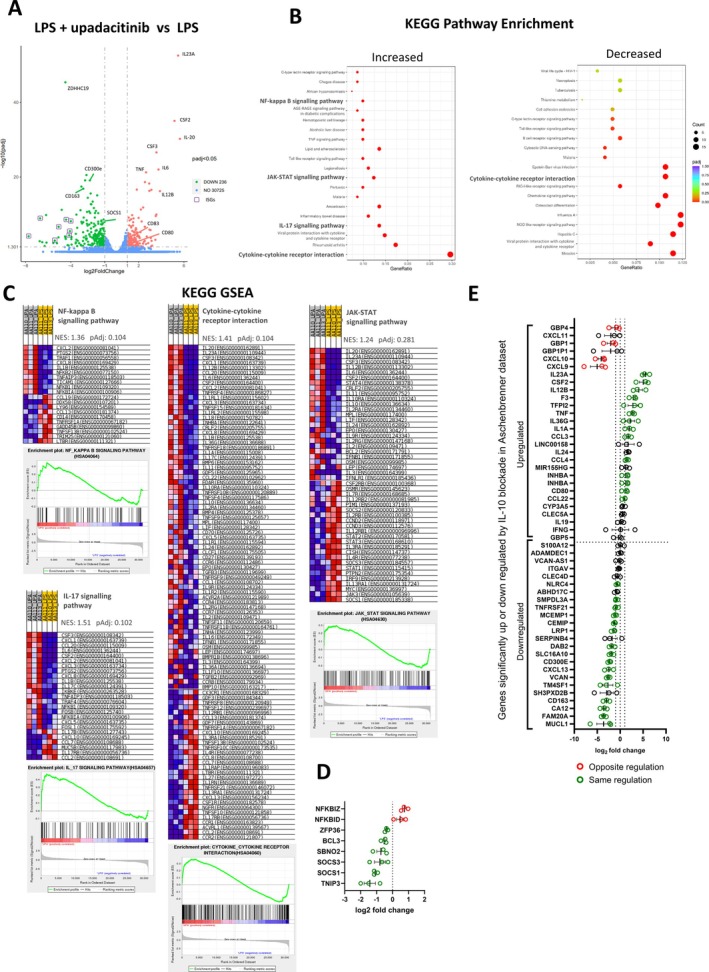
Methods: Spinal entheseal tissue was activated with Toll-like receptor (TLR) agonists, including TLR4 and interleukin-1 (IL-1) family proteins, and entheseal T cells were activated with anti-CD3/anti-CD28 with IL-23/IL-1β. JAKi via upadacitinib (JAK1/JAK2), deucravacitinib (Tyk2), and ritlecitinib (JAK3) inhibition was evaluated using multiplex cytokine assays, intracellular flow, and bulk RNA sequencing (RNAseq) and cytokine blocking or stimulation.
Results: Following interferon γ stimulation, JAK1 inhibition blocked STAT1 phosphorylation in entheseal cells and strongly blocked activated entheseal T cell tumor necrosis factor α (TNFα), IL-17A, and IL-17F production. The opposite effect was evident in entheseal myeloid cell with exaggerated TLR4 and other adjuvant-mediated cytokine production including IL-23 (~10-fold increase; P < 0.001) or TNFα (~10-fold increase; P < 0.0001). This myeloid effect was induced by upadacitinib and deucravacitinib but not ritlecitinib, suggesting IL-10R JAK1/Tyk2 signaling. Bulk RNAseq showed a multifaceted impact of JAKi on myeloid activation with strong M1 type monocyte polarization under TLR4 stimulation and JAK1 inhibition confirmed by flow cytometry. Direct IL-10 inhibition recapitulated inflammatory cytokine elevations and IL-10R agonist largely, but not completely, rescued this phenotype.
Conclusion: These findings help explain the emergent efficacy of Tyk2 blockade in SpA spectrum-related arthritis that is not IL-10 dependent but indicates why such strategies may not be a panacea for SpA spectrum disorder-related intestinal inflammation.
© 2025 The Author(s). Arthritis & Rheumatology published by Wiley Periodicals LLC on behalf of American College of Rheumatology.
Figures





References
-
- Sherlock JP, Joyce‐Shaikh B, Turner SP, et al. IL‐23 induces spondyloarthropathy by acting on ROR‐γt+ CD3+CD4−CD8− entheseal resident T cells. Nat Med 2012;18(7):1069–1076. - PubMed
-
- Bridgewood C, Sharif K, Sherlock J, et al. Interleukin‐23 pathway at the enthesis: the emerging story of enthesitis in spondyloarthropathy. Immunol Rev 2020;294(1):27–47. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
