

Transient DREADD Manipulation of the Dorsal Dentate Gyrus in Rats Impairs Initial Learning of Place-Outcome Associations
- PMID: 40326013
- PMCID: PMC12053149
- DOI: 10.1002/hipo.70014
Transient DREADD Manipulation of the Dorsal Dentate Gyrus in Rats Impairs Initial Learning of Place-Outcome Associations
Abstract
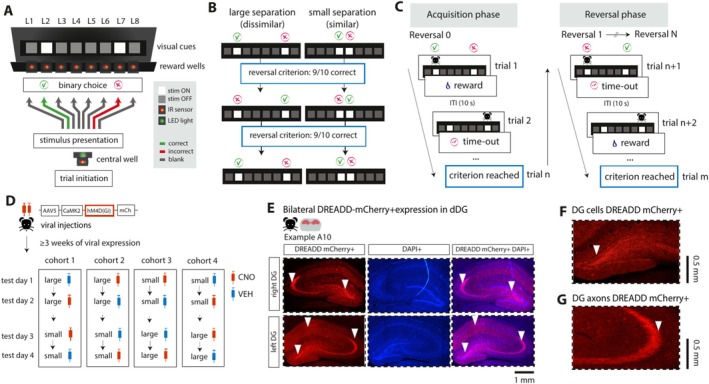
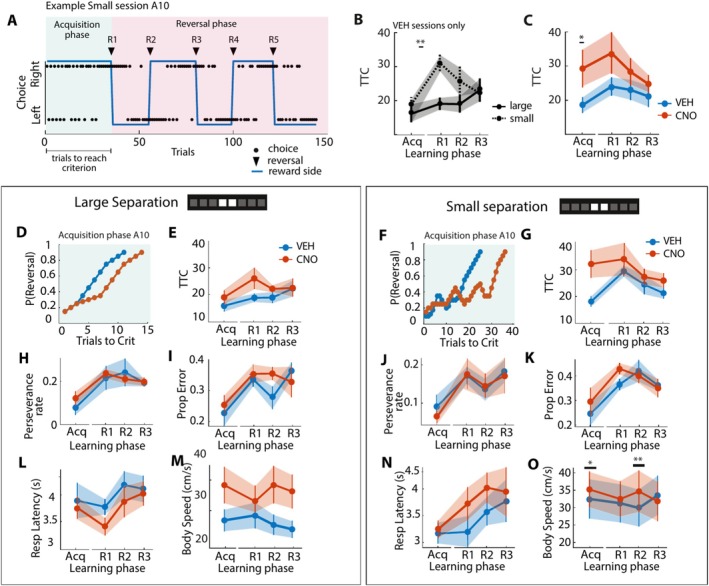
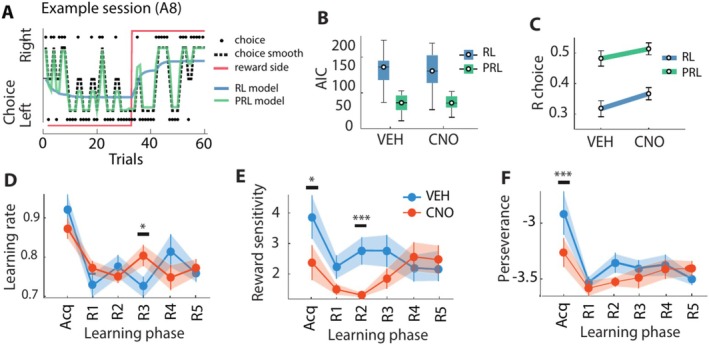
The dentate gyrus subfield of the hippocampus is thought to be critically involved in the disambiguation of similar episodic experiences and places in a context-dependent manner. However, most empirical evidence has come from lesion and gene knock-out studies in rodents, in which the dentate gyrus is permanently perturbed and compensation of affected functions via other areas within the memory circuit could take place. The acute and causal role of the dentate gyrus herein remains therefore elusive. The present study aimed to investigate the acute role of the dorsal dentate gyrus in disambiguation learning using reversible inhibitory DREADDs. Rats were trained on a location discrimination task and learned to discriminate between a rewarded and unrewarded location with either small (similar condition) or large (dissimilar condition) separation. Reward contingencies switched after applying a reversal rule, allowing us to track the temporal engagement of the dentate gyrus during the task. Bilateral DREADD modulation of the dentate gyrus impaired the initial acquisition learning of place-reward associations, but performance rapidly recovered to baseline levels within the same session. Modeling of the behavioral patterns revealed that reward sensitivity and alternation behavior were temporally associated with the DG-dependent impairment during acquisition learning. Our study thus provides novel evidence that the dorsal dentate gyrus is acutely engaged during the initial acquisition learning of place-reward associations.
Keywords: DREADDs; dentate gyrus; episodic memory; hippocampus; place‐reward associations; rats.
© 2025 The Author(s). Hippocampus published by Wiley Periodicals LLC.
Figures
References
-
- Akaike, H. 1998. “Information Theory and an Extension of the Maximum Likelihood Principle.” In Selected Papers of Hirotugu Akaike, edited by Parzen E., Tanabe K., and Kitagawa G., 199–213. Springer.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
