

Functional role of cell classes in monkey prefrontal cortex after learning a working memory task
- PMID: 40328904
- PMCID: PMC12056203
- DOI: 10.1038/s42003-025-08142-4
Functional role of cell classes in monkey prefrontal cortex after learning a working memory task
Abstract
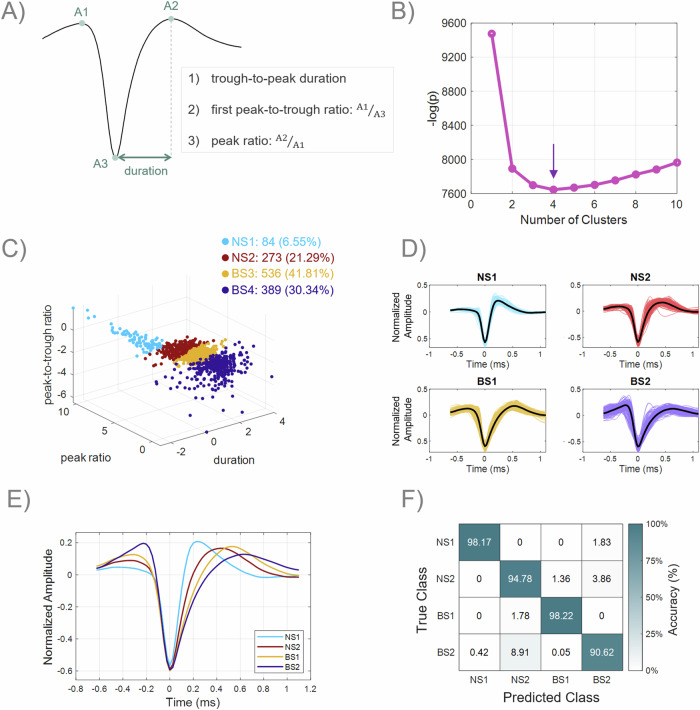
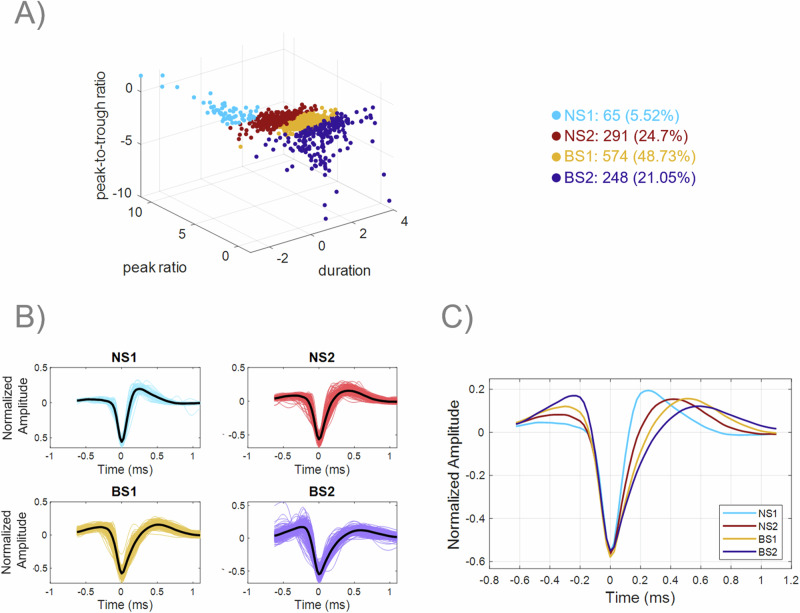
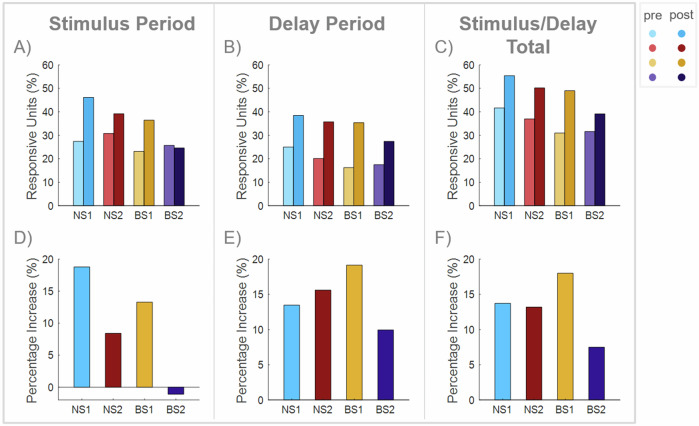
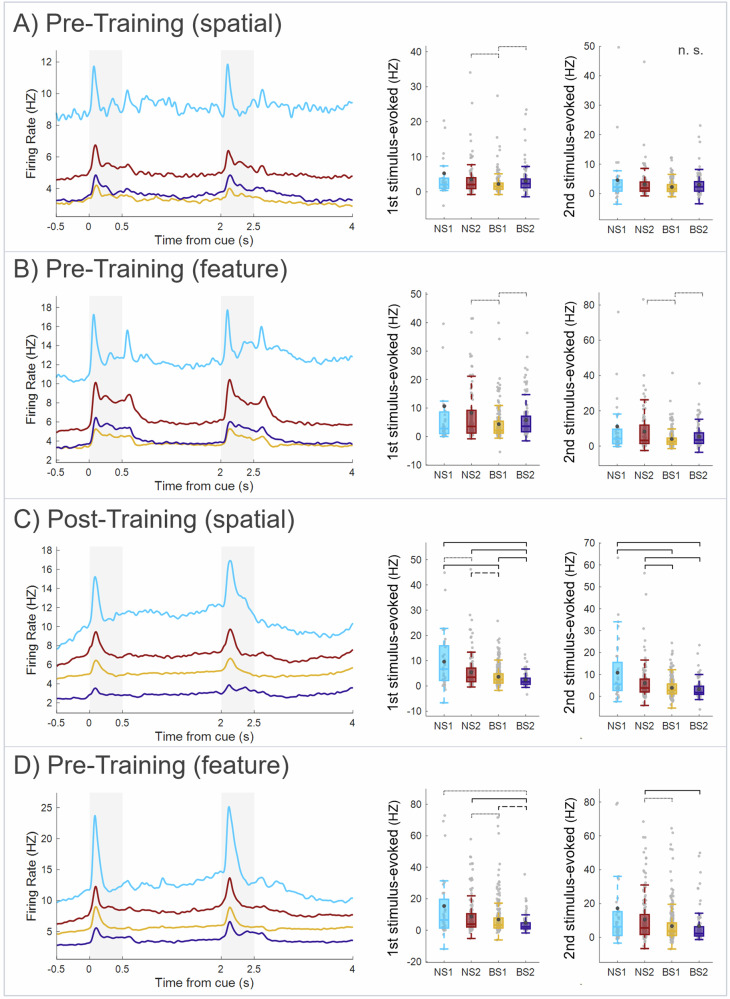
The prefrontal cortex (PFC) is important for learning and performing working memory tasks. However, its precise role for spatial and non-spatial working memory, and the role of different cell types in the circuits that maintain working memory remain poorly understood. To investigate this issue, we analyzed single-unit recordings from the PFC of monkeys during the passive viewing phase before they learned the task rules and after learning, during the execution of active working memory tasks (spatial and feature). Through cluster analysis of extracellular spike waveform features, we identified two classes of narrow-spiking neurons (putative inhibitory cells) and two classes of broad-spiking neurons (putative pyramidal cells). These putative cell classes exhibited distinct physiological characteristics, including baseline firing rates, baseline neural firing variability, and visual stimulus-evoked responses. Neuronal response modulation varied heterogeneously across these cell classes after training and performing active tasks. Training and execution of spatial working memory resulted in higher activity in all class types, highlighting the involvement of diverse prefrontal circuits in spatial information processing. In contrast, feature working memory training and execution affected activity of broad-spiking cell classes alone, suggesting less involvement of a prefrontal circuit in the representation of feature information. We also revealed hitherto unknown, differential effects of training and task execution on different broad-spiking cell types. One broad-spiking neuron subtype exhibited significant response modulation, with increased baseline firing rate, stimulus-evoked responses, and working memory-related firing rates. Another broad-spiking subtype showed decreased baseline firing rate and variability, which may optimize neural coding efficiency. This study advances our understanding of the functional heterogeneity within the PFC and the specialized contributions of different neuronal subtypes to cognitive processes.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


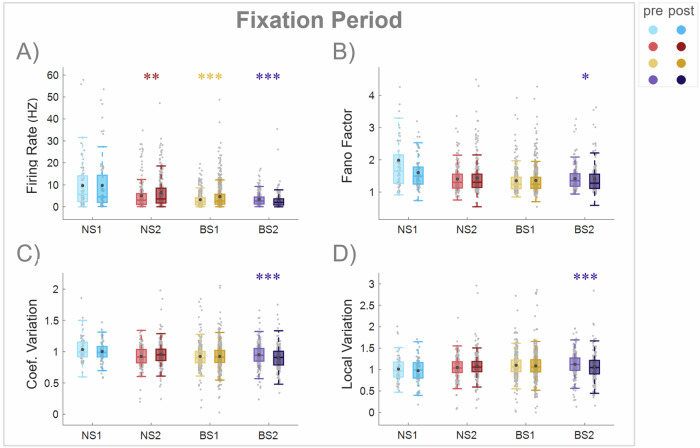



Similar articles
-
Changes in prefrontal neuronal activity after learning to perform a spatial working memory task.Cereb Cortex. 2011 Dec;21(12):2722-32. doi: 10.1093/cercor/bhr058. Epub 2011 Apr 28. Cereb Cortex. 2011. PMID: 21527786 Free PMC article.
-
Trial-to-Trial Variability of Spiking Delay Activity in Prefrontal Cortex Constrains Burst-Coding Models of Working Memory.J Neurosci. 2021 Oct 27;41(43):8928-8945. doi: 10.1523/JNEUROSCI.0167-21.2021. Epub 2021 Sep 22. J Neurosci. 2021. PMID: 34551937 Free PMC article.
-
Emergence of Nonlinear Mixed Selectivity in Prefrontal Cortex after Training.J Neurosci. 2021 Sep 1;41(35):7420-7434. doi: 10.1523/JNEUROSCI.2814-20.2021. Epub 2021 Jul 22. J Neurosci. 2021. PMID: 34301827 Free PMC article.
-
Diverse synchrony of firing reflects diverse cell-assembly coding in the prefrontal cortex.J Physiol Paris. 2013 Dec;107(6):459-70. doi: 10.1016/j.jphysparis.2013.05.004. Epub 2013 Jun 4. J Physiol Paris. 2013. PMID: 23747709 Review.
-
Plasticity of Persistent Activity and Its Constraints.Front Neural Circuits. 2020 May 7;14:15. doi: 10.3389/fncir.2020.00015. eCollection 2020. Front Neural Circuits. 2020. PMID: 32528254 Free PMC article. Review.
References
-
- Baddeley, A. Working memory. Science255, 556–559 (1992). - PubMed
-
- Gathercole, S. E., Brown, L. & Pickering, S. J. Working memory assessments at school entry as longitudinal predictors of National Curriculum attainment levels. Educ. Child Psychol.20, 109–122 (2003).
-
- Baddeley, A. Working memory: Theories, models, and controversies. Annu. Rev. Psychol.63, 1–29 (2012). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
