

Effect of RNA on the supramolecular architecture of α-synuclein fibrils
- PMID: 40329536
- PMCID: PMC12256921
- DOI: 10.1016/j.bpj.2025.04.031
Effect of RNA on the supramolecular architecture of α-synuclein fibrils
Abstract
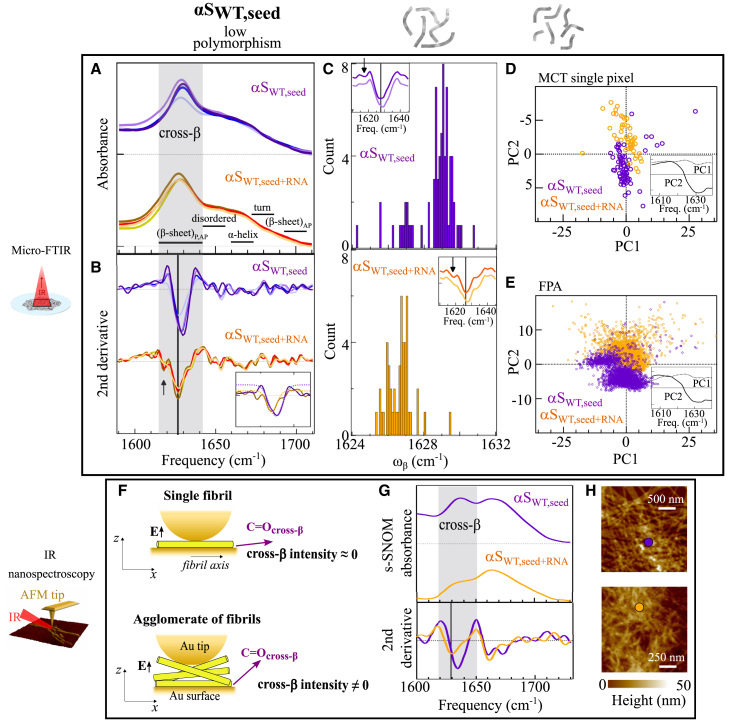
Structural changes associated with protein aggregation are challenging to study, requiring the combination of experimental techniques providing insights at the molecular level across diverse scales, ranging from nanometers to microns. Understanding these changes is even more complex when aggregation occurs in the presence of molecular cofactors such as nucleic acids and when the resulting aggregates are highly polymorphic. Infrared (IR) spectroscopy is a powerful tool for studying protein aggregates since it combines the label-free sensitivity to the cross-β architecture, an inherent feature of protein supramolecular aggregates, with the possibility to reach nanoscale sensitivity by leveraging atomic force microscopy (AFM)-assisted detection. Here, we present a combined approach that detects IR spectral markers of aggregation using various IR spectroscopy techniques, covering micro-to-nanoscale ranges, to study the effect of RNA on the supramolecular architecture of α-synuclein amyloid aggregates. We show a clear impact of RNA consistent with enhanced intermolecular forces, likely via a stronger hydrogen-bonded network stabilizing the cross-β architecture. AFM-assisted IR spectroscopy was crucial to assess that the more ordered the aggregates are, the stronger the structural impact of RNA. In addition, an RNA-induced reduction of the degree of polymorphism within the aggregate population is obtained.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
-
- Louros N., Schymkowitz J., Rousseau F. Mechanisms and pathology of protein misfolding and aggregation. Nat. Rev. Mol. Cell Biol. 2023;24:912–933. - PubMed
-
- Vekrellis K., Xilouri M., et al. Stefanis L. Pathological roles of α-synuclein in neurological disorders. Lancet Neurol. 2011;10:1015–1025. - PubMed
-
- Galvin J.E., Lee V.M., Trojanowski J.Q. Synucleinopathies: clinical and pathological implications. Arch. Neurol. 2001;58:186–190. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
