

Lumacaftor inhibits channel activity of rescued F508del cystic fibrosis transmembrane conductance regulator
- PMID: 40331529
- PMCID: PMC12173061
- DOI: 10.1152/ajplung.00287.2024
Lumacaftor inhibits channel activity of rescued F508del cystic fibrosis transmembrane conductance regulator
Abstract
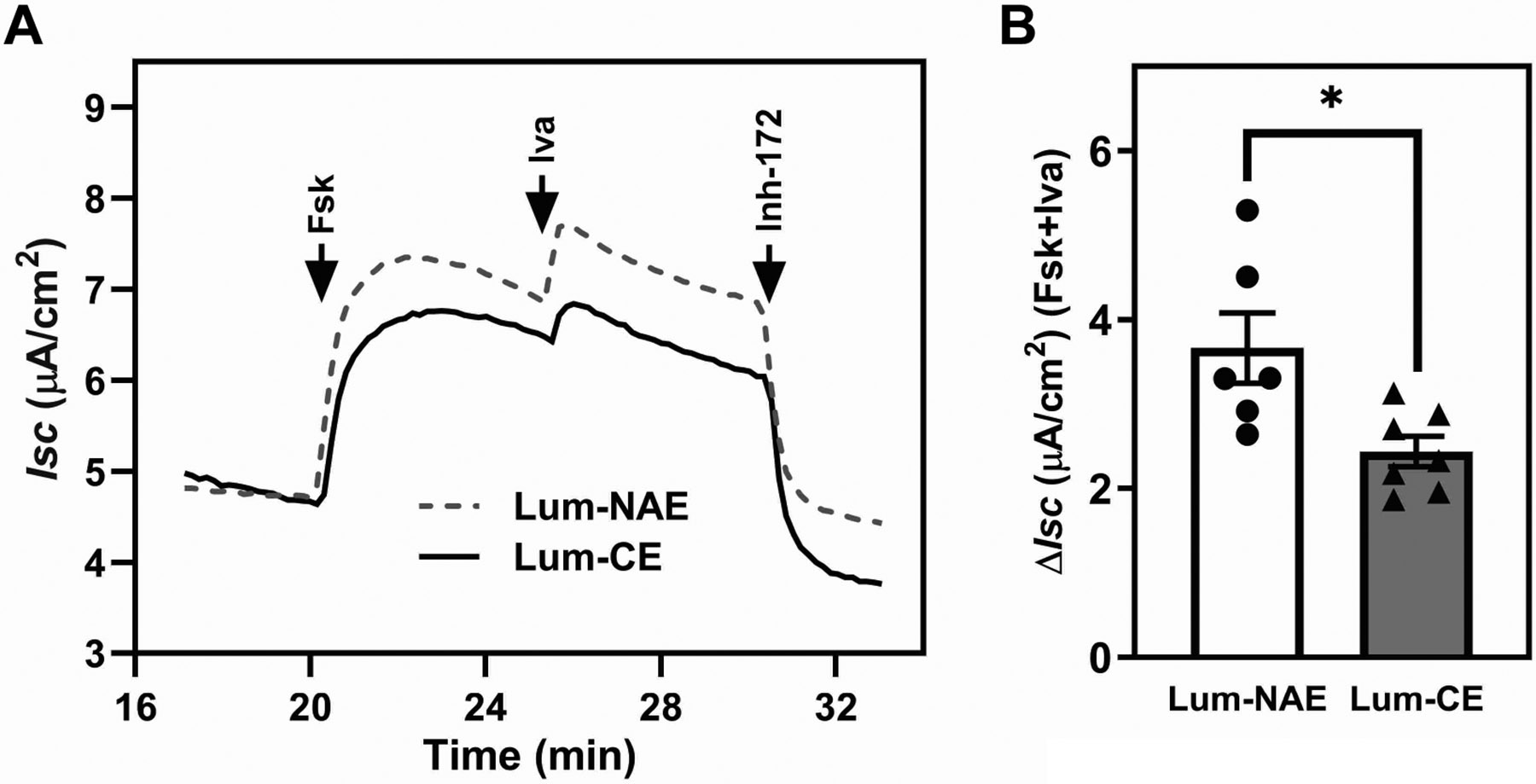
Lumacaftor, the corrector of Orkambi, enhances the processing of F508del cystic fibrosis transmembrane conductance regulator (CFTR), but its impact on the channel activity of rescued F508del CFTR (rF508del) is unclear. Using an electrode-based, real-time iodide efflux assay performed at room temperature, acute exposure to lumacaftor was shown to increase the processing of F508del CFTR without a proportional increase in channel activity in a CFBE41o-cell line stably expressing F508del CFTR (CFBE-DF). A similar effect was not observed on wild-type CFTR in a HEK293 cell line. At 37°C, rF508del channel activity is significantly inhibited in CFBE-DF cells by acute exposure to 5 µM lumacaftor, but not to 5 µM tezacaftor or 1 µM elexacaftor, the two correctors of Trikafta. Lumacaftor's inhibitory effect was characterized by a major left shift of the peak channel activity relative to the peak CFTR processing in the dose-response chart, which is absent for tezacaftor or elexacaftor. Ussing chamber analysis on polarized CFBE-DF cells reveals an inhibitory effect for lumacaftor on the forskolin- and ivacaftor-induced change in short-circuit current. Single channel patch clamp on HEK-DF cells shows that acute application of cytosolic lumacaftor significantly decreases rF508del channel open probability. Taken together, despite its strong corrector activity, lumacaftor inhibits rF508del channel activity, compromising the degree of functional rescue. This effect may contribute to the limited clinical efficacy of Orkambi.NEW & NOTEWORTHY Small-molecule correctors bind to F508del cystic fibrosis transmembrane conductance regulator (CFTR) and restore its trafficking to the plasma membrane to function as an anion channel. Despite its high efficacy as a corrector, lumacaftor inhibits the channel opening of rescued F508del CFTR, making it a weak CFTR modulator. The current work highlights the impact of CFTR correctors on the channel activity of rescued F508del CFTR as an important variable in the efficacy of modulator therapy.
Keywords: cystic fibrosis; cystic fibrosis transmembrane conductance regulator (CFTR); iodide efflux assay; ion channel; lumacaftor.
Figures








References
-
- Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL, Drumm ML, Iannuzzi MC, Collins FS, Tsui L-C. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245: 1066–73, 1989. - PubMed
-
- Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW, White GA, O’Riordan CR, Smith AE. Defective intracellular transport and processing of CFTR is the molecular basis of most cystic fibrosis. Cell 63: 827–34, 1990. - PubMed
-
- Du K, Sharma M, Lukacs GL. The DeltaF508 cystic fibrosis mutation impairs domain-domain interactions and arrests post-translational folding of CFTR. Nat Struct Mol Biol 12: 17–25, 2005. - PubMed
-
- Dalemans W, Barbry P, Champigny G, Jallat S, Dott K, Dreyer D, Crystal RG, Pavirani A, Lecocq JP, Lazdunski M. Altered chloride ion channel kinetics associated with the delta F508 cystic fibrosis mutation. Nature 354: 526–8, 1991. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous