

The structure of the Vibrio natriegens 70S ribosome in complex with the proline-rich antimicrobial peptide Bac5(1-17)
- PMID: 40331629
- PMCID: PMC12056610
- DOI: 10.1093/nar/gkaf324
The structure of the Vibrio natriegens 70S ribosome in complex with the proline-rich antimicrobial peptide Bac5(1-17)
Abstract
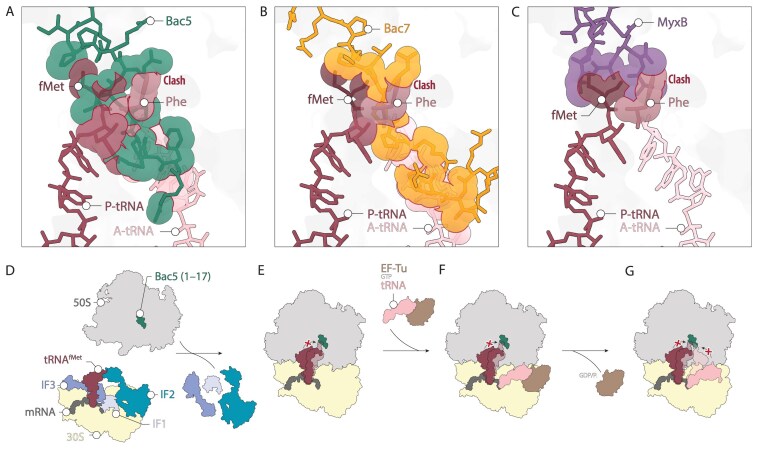
Proline-rich antimicrobial peptides (PrAMPs) are produced as part of the innate immune response of animals, insects, and plants. The well-characterized mammalian PrAMP bactenecin-5 (Bac5) has been shown to help fight bacterial infection by binding to the bacterial ribosome and inhibiting protein synthesis. In the absence of Bac5-ribosome structures, the binding mode of Bac5 and exact mechanism of action has remained unclear. Here, we present a cryo-electron microscopy structure of Bac5 in complex with the 70S ribosome from the Gram-negative marine bacterium Vibrio natriegens. The structure shows that, despite sequence similarity to Bac7 and other type I PrAMPs, Bac5 displays a completely distinct mode of interaction with the ribosomal exit tunnel. Bac5 overlaps with the binding site of both A- and P-site transfer RNAs bound at the peptidyltransferase center, suggesting that this type I PrAMP can interfere with late stages of translation initiation as well as early stages of elongation. Collectively, our study presents a ribosome structure from V. natriegens, a fast-growing bacterium that has interesting biotechnological and synthetic biology applications, as well as providing additional insights into the diverse binding modes that type I PrAMPs can utilize to inhibit protein synthesis.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
None declared.
Figures






References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
