

Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice
- PMID: 40335415
- PMCID: PMC12173048
- DOI: 10.1016/j.ebiom.2025.105734
Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice
Abstract
Background: Tyrosine protein-kinase 2 (TYK2) mediates inflammatory signalling through multiple cytokines, including interferon-α (IFNα), interleukin (IL)-12, and IL-23. TYK2 missense mutations protect against type 1 diabetes (T1D), and inhibition of TYK2 shows promise in other autoimmune conditions.
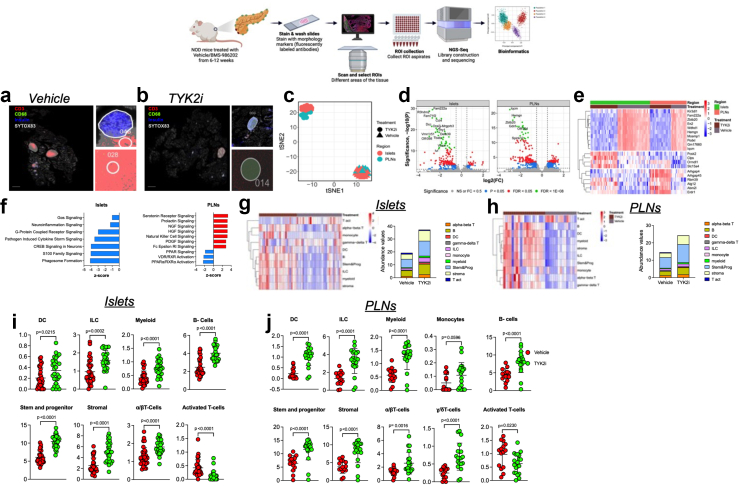
Methods: We evaluated the effects of specific TYK2 inhibitors (TYK2is) in pre-clinical models of T1D, including human β cells, cadaveric islets, iPSC-derived islets, and mouse models.
Findings: In vitro studies showed that TYK2is prevented IFNα-induced β cell HLA class I up-regulation, endoplasmic reticulum stress, and chemokine production. In co-culture studies, pre-treatment of β cells with TYK2i prevented IFNα-induced antigenic peptide presentation and alloreactive and autoreactive T cell degranulation. In vivo administration of BMS-986202 in two mouse models of T1D (RIP-LCMV-GP and NOD mice) reduced systemic and tissue-localised inflammation, prevented β cell death, and delayed T1D onset. Transcriptional phenotyping of pancreatic islets, pancreatic lymph nodes, and spleen highlighted a role for TYK2 inhibition in modulating signalling pathways associated with inflammation, translational control, stress signalling, secretory function, immunity, and diabetes. Additionally, TYK2i treatment changed the composition of innate and adaptive immune cell populations in the blood and disease target tissues.
Interpretation: These findings indicate that TYK2i has beneficial effects on both the immune and endocrine compartments in models of T1D, thus supporting a path forward for testing TYK2is in human T1D.
Funding: This work was supported by the National Institutes of Health (NIH), Veteran Affairs (VA), Breakthrough T1D, and gifts from the Sigma Beta Sorority, the Ball Brothers Foundation, and the George and Frances Ball Foundation.
Keywords: Interferon-α; Islets of langerhans; T cell; Type 1 diabetes; Tyrosine protein-kinase 2 (TYK2); β cell.
Published by Elsevier B.V.
Conflict of interest statement
Declaration of interests CEM has received grants from Lilly Pharmaceuticals and Astellas Pharmaceuticals (not related to this manuscript). CEM has served on advisory boards related to T1D research clinical trial initiatives: Isla Technologies, Neurodon, and DiogenX. CEM has patent (16/291,668) Extracellular Vesicle Ribonucleic Acid (RNA) Cargo as a Biomarker of Hyperglycaemia and Type 1 Diabetes and CEM and FS have a provisional patent (63/285,765) Biomarker for Type 1 Diabetes (PDIA1 as a biomarker of β cell stress). DLE serves on the advisory board of InSphero, related to the preparation of islet microtissues. EMV received grant from Fonds de la Recherche. JSK has received grants from NIDDK—U24DK104162, R21DK127285 and grants from (Breakthrough T1D–5-SRA-2018-557-Q-R) and Helmsley Charitable Trust–2018 PG-T1D053 and G-2108-04793). JSK received support to attend NIDDK study section, as well as to attend the NIDDK sponsored AI meeting October 2024. SAW is employed by Eli Lilly and owns stocks in the company and was supported by the company to attend a conference. SAW is a board member of Breakthrough T1D. FS received honoria and a travel award to present current work at ADA conference. These activities have not dealt directly with topics covered in this manuscript.
Figures







Update of
-
Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice.bioRxiv [Preprint]. 2024 May 9:2024.03.20.585925. doi: 10.1101/2024.03.20.585925. bioRxiv. 2024. Update in: EBioMedicine. 2025 Jul;117:105734. doi: 10.1016/j.ebiom.2025.105734. PMID: 38766166 Free PMC article. Updated. Preprint.
References
-
- Platanias L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat Rev Immunol. 2005;5(5):375–386. - PubMed
-
- Nyaga D.M., Vickers M.H., Jefferies C., Perry J.K., O'Sullivan J.M. Type 1 diabetes mellitus-associated genetic variants contribute to overlapping immune regulatory networks. Front Genet. 2018;9 https://www.frontiersin.org/articles/10.3389/fgene.2018.00535 [cited 2023 Aug 12]. Available from: - DOI - PMC - PubMed
-
- Tao J.H., Zou Y.F., Feng X.L., et al. Meta-analysis of TYK2 gene polymorphisms association with susceptibility to autoimmune and inflammatory diseases. Mol Biol Rep. 2011;38(7):4663–4672. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
