

Pancreatic cancer-restricted cryptic antigens are targets for T cell recognition
- PMID: 40339010
- PMCID: PMC12163983
- DOI: 10.1126/science.adk3487
Pancreatic cancer-restricted cryptic antigens are targets for T cell recognition
Abstract
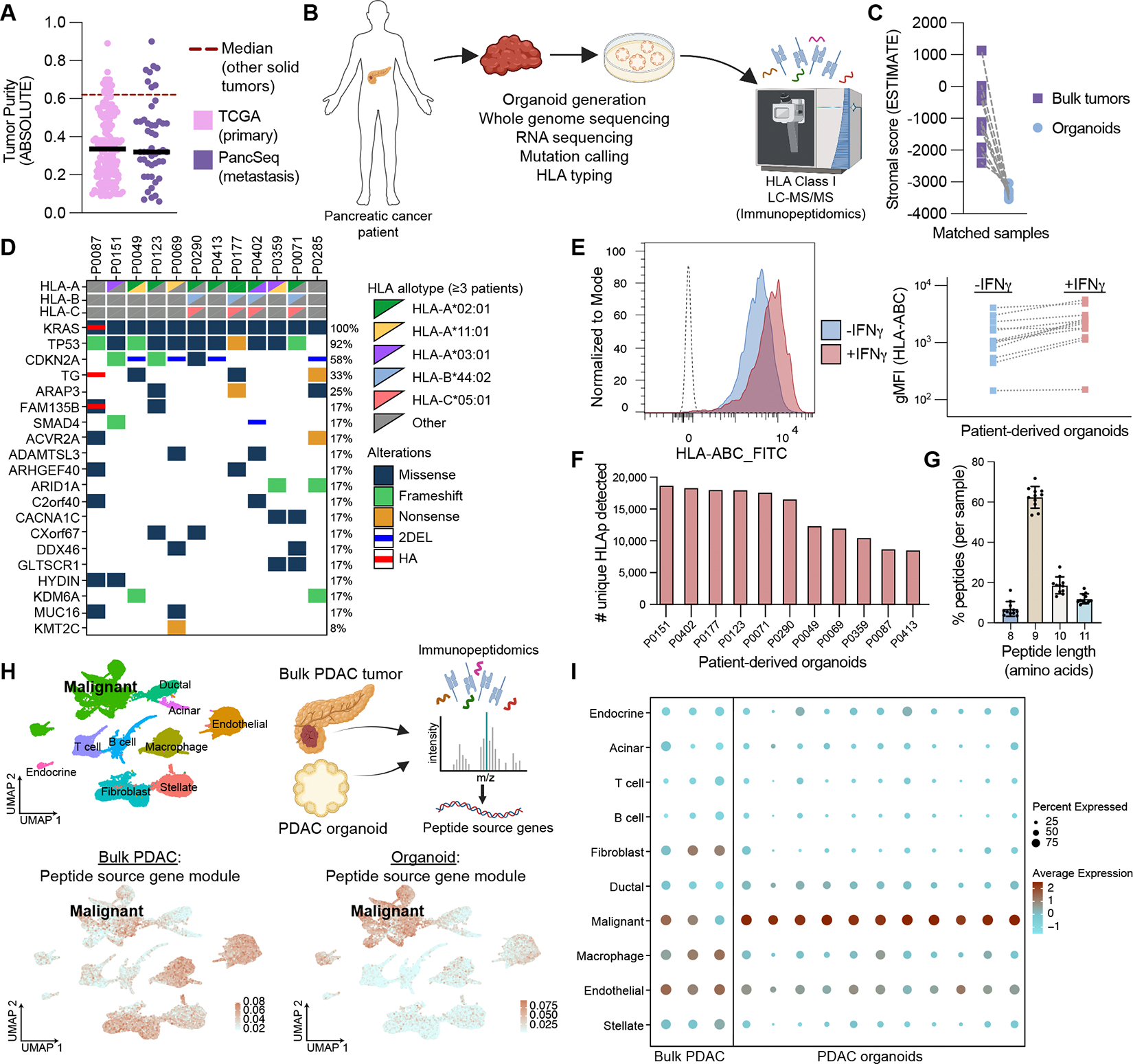
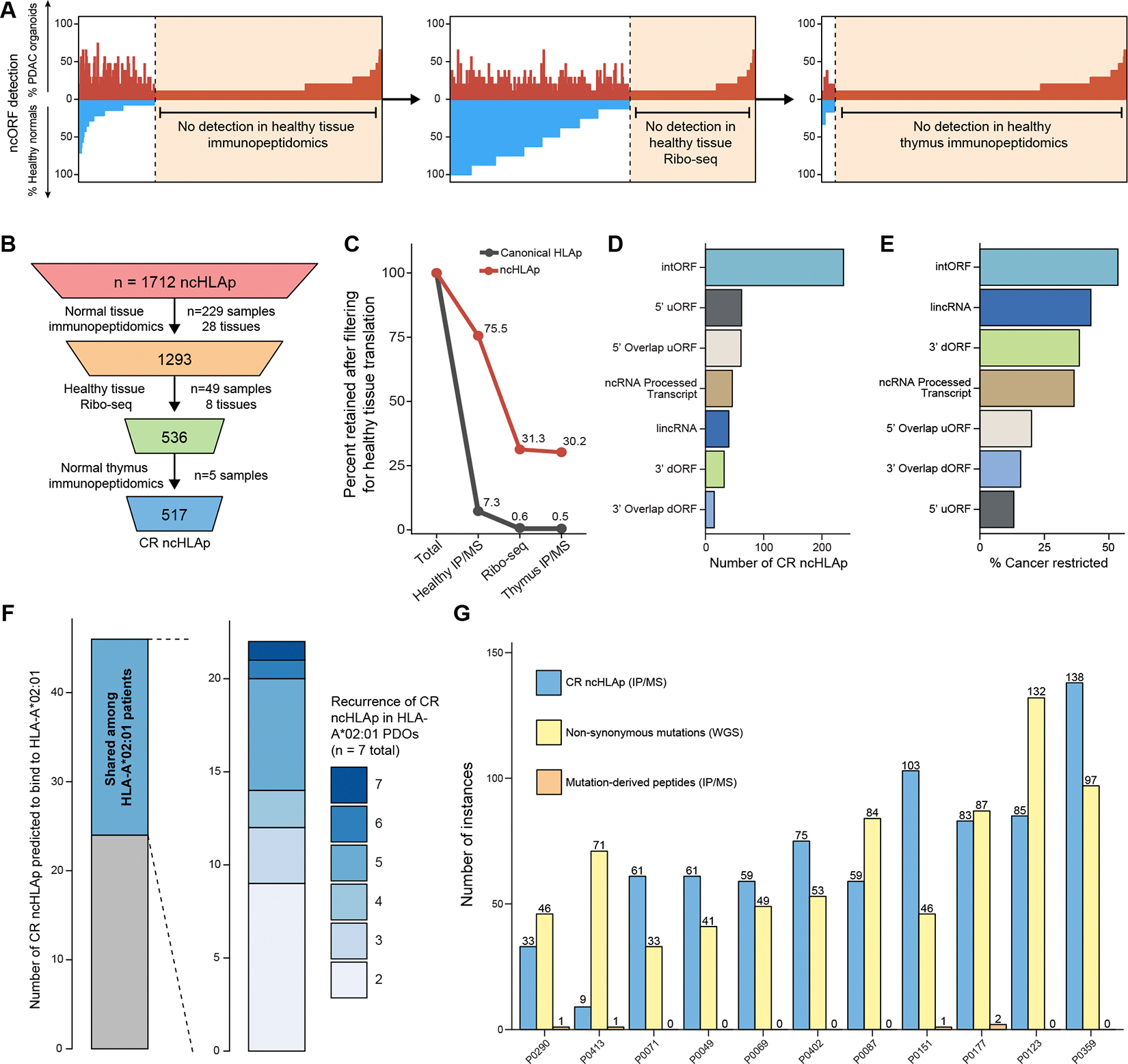
Translation of the noncoding genome in cancer can generate cryptic (noncanonical) peptides capable of presentation by human leukocyte antigen class I (HLA-I); however, the cancer specificity and immunogenicity of noncanonical HLA-I-bound peptides (ncHLAp) are incompletely understood. Using high-resolution immunopeptidomics, we discovered that cryptic peptides are abundant in the pancreatic cancer immunopeptidome. Approximately 30% of ncHLAp exhibited cancer-restricted translation, and a substantial subset were shared among patients. Cancer-restricted ncHLAp displayed robust immunogenic potential in a sensitive ex vivo T cell priming platform. ncHLAp-reactive, T cell receptor-redirected T cells exhibited tumoricidal activity against patient-derived pancreatic cancer organoids. These findings demonstrate that pancreatic cancer harbors cancer-restricted ncHLAp that can be recognized by cytotoxic T cells. Future therapeutic strategies for pancreatic cancer, and potentially other solid tumors, may include targeting cryptic antigens.
Conflict of interest statement
Figures



Comment in
-
The hunt for common tumor antigens.Science. 2025 May 8;388(6747):592-593. doi: 10.1126/science.adx8688. Epub 2025 May 8. Science. 2025. PMID: 40339036
References
-
- Chong C, Müller M, Pak HS, Harnett D, Huber F, Grun D, Leleu M, Auger A, Arnaud M, Stevenson BJ, Michaux J, Bilic I, Hirsekorn A, Calviello L, Simó-Riudalbas L, Planet E, Lubiński J, Bryśkiewicz M, Wiznerowicz M, Xenarios I, Zhang L, Trono D, Harari A, Ohler U, Coukos G, Bassani-Sternberg M, Integrated proteogenomic deep sequencing and analytics accurately identify non-canonical peptides in tumor immunopeptidomes. Nat Commun 11, 1293 (2020). - PMC - PubMed
-
- Laumont CM, Vincent K, Hesnard L, Audemard É, Bonneil É, Laverdure JP, Gendron P, Courcelles M, Hardy MP, Côté C, Durette C, St-Pierre C, Benhammadi M, Lanoix J, Vobecky S, Haddad E, Lemieux S, Thibault P, Perreault C, Noncoding regions are the main source of targetable tumor-specific antigens. Sci Transl Med 10 (2018). - PubMed
-
- Lozano-Rabella M, Garcia-Garijo A, Palomero J, Yuste-Estevanez A, Erhard F, Farriol-Duran R, Martín-Liberal J, Ochoa-De-Olza M, Matos I, Gartner JJ, Ghosh M, Canals F, Vidal A, Piulats JM, Matías-Guiu X, Brana I, Mu~ Noz-Couselo E, Garralda E, Schlosser A, Gros A, Exploring the Immunogenicity of Noncanonical HLA-I Tumor Ligands Identified through Proteogenomics. Clinical Cancer Research 29, 2250–2265 (2023). - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
