

Modulation of antibody transport in the brain and spinal cord through the intranasal pathway
- PMID: 40340136
- PMCID: PMC12418472
- DOI: 10.1016/j.neurot.2025.e00606
Modulation of antibody transport in the brain and spinal cord through the intranasal pathway
Abstract
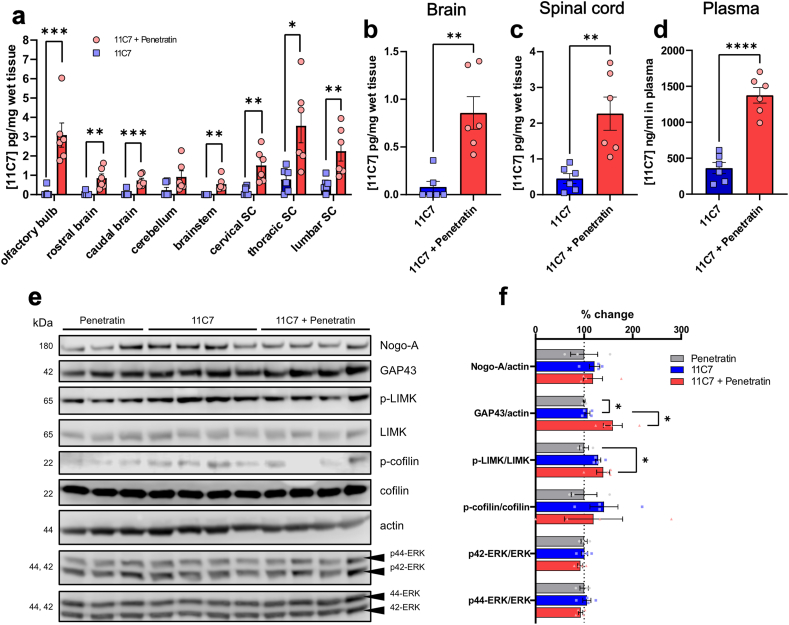
The intranasal pathway is a promising antibody delivery route for the treatment of neurological diseases, but the mechanisms mediating nose-to-brain/spinal cord transport are poorly understood. The aim of our study was to determine if the transport of antibodies can pharmacologically be modulated in the mouse CNS. The pharmacokinetics and distribution of recombinant antibodies were followed in brain and spinal cord homogenates and biofluids by ELISA and immunofluorescence. A non-CNS antigen-binding antibody (FG12) was used to monitor target-independent transport whereas 11C7 mAb, neutralizing the myelin-associated growth inhibitor Nogo-A, was applied to induce CNS target-dependent neuronal growth response. Fast axonal transport/neuronal activity were inhibited with Lidocaine pre-treatment on the olfactory mucosa. Antibody uptake was enhanced across the olfactory epithelium with the co-administration of the cell-penetrating peptide Penetratin. Growth signalling pathways were examined by Western blotting. FG12 was detected in the brain and spinal cord as early as 30 min after intranasal administration. After 1 h, the concentration of FG12 rapidly declined in all CNS areas and was back to baseline values at 24 h. Lidocaine prevented the early rise in FG12 concentration in the spinal cord. This effect was not observed in the brain. Penetratin allowed to maintain the elevation of FG12 and to activate 11C7-induced growth signalling in the spinal cord at 24 h. Our data suggest that the pharmacological modulation of transport mechanisms in the nose-to-CNS pathways may allow to control the therapeutic effects of antibodies in neurological diseases.
Keywords: Blood-brain barrier; Cell-penetrating peptide; Intranasal pathway; Neuronal plasticity; Nogo-A; Recombinant antibodies.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest AC has received speakers’/board honoraria from Actelion (Janssen/J&J), Almirall, Bayer, Biogen, Celgene (BMS), Genzyme, Merck KGaA (Darmstadt, Germany), Novartis, Roche, and Teva, all for hospital research funds. He received research support from Biogen, Genzyme, and UCB, the European Union, and the Swiss National Foundation. He serves as associate editor of the European Journal of Neurology, on the editorial board for Clinical and Translational Neuroscience and as topic editor for the Journal of International Medical Research.
Figures








References
-
- Shen J., Attar M. Antibody-Drug conjugate (ADC) research in ophthalmology--a review. Pharm Res. 2015;32(11):3572–3576. - PubMed
-
- Nisenbaum L., Martone R., Chen T., Rajagovindan R., Dent G., Beaver J., et al. CSF biomarker concordance with amyloid PET in Phase 3 studies of aducanumab. Alzheimer’s Dementia : the journal of the Alzheimer’s Association. 2023;19(8):3379–3388. - PubMed
-
- Perneczky R., Jessen F., Grimmer T., Levin J., Flöel A., Peters O., et al. Anti-amyloid antibody therapies in Alzheimer’s disease. Brain : J Neurol. 2023;146(3):842–849. - PubMed
-
- Winkelmann A., Loebermann M., Reisinger E.C., Hartung H.-P., Zettl U.K. Disease-modifying therapies and infectious risks in multiple sclerosis. 2016;12(4):217–233. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
