

Control of replication and gene expression by ADP-ribosylation of DNA in Mycobacterium tuberculosis
- PMID: 40341764
- PMCID: PMC12170906
- DOI: 10.1038/s44318-025-00451-y
Control of replication and gene expression by ADP-ribosylation of DNA in Mycobacterium tuberculosis
Erratum in
-
Author Correction: Control of replication and gene expression by ADP-ribosylation of DNA in Mycobacterium tuberculosis.EMBO J. 2025 Aug;44(15):4406. doi: 10.1038/s44318-025-00491-4. EMBO J. 2025. PMID: 40562877 Free PMC article.
Abstract
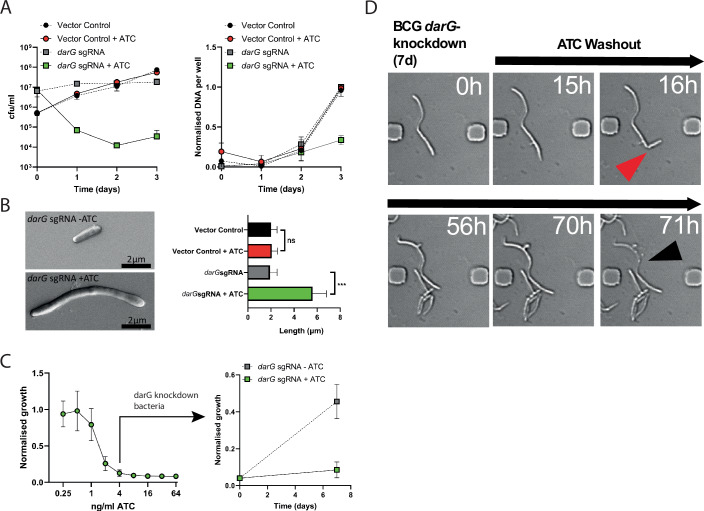
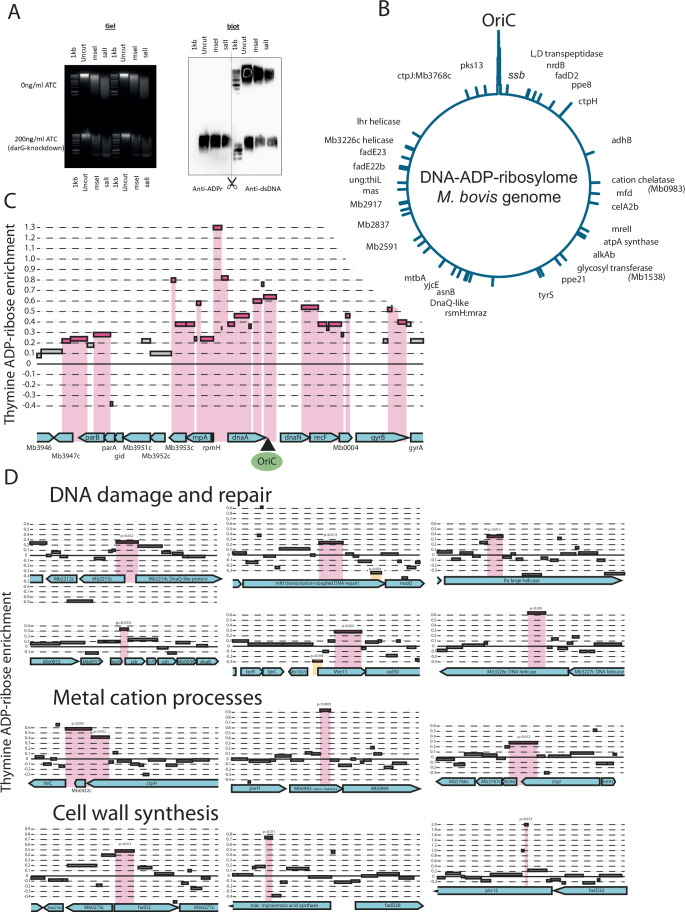
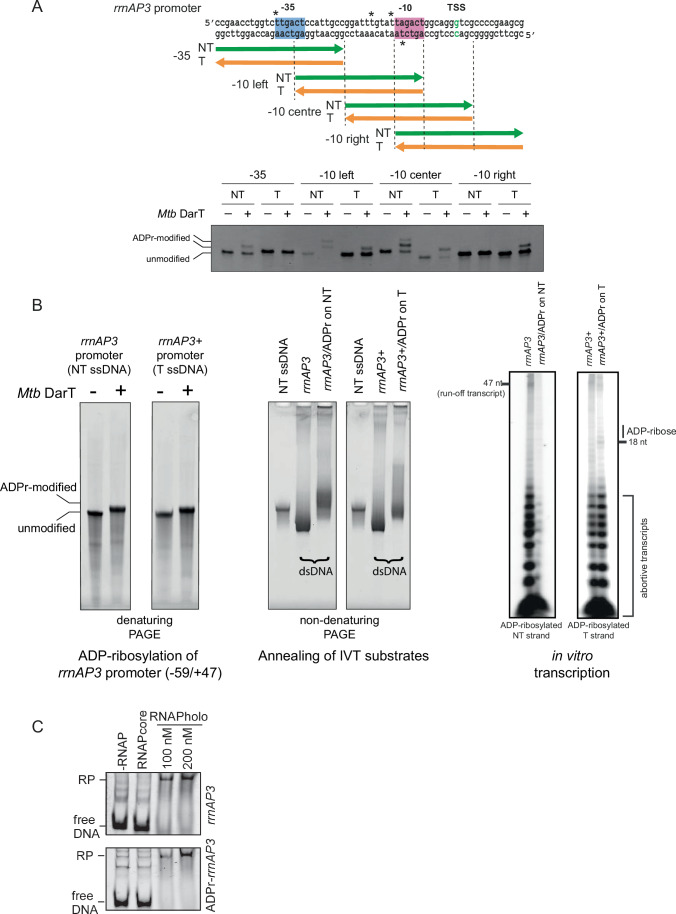
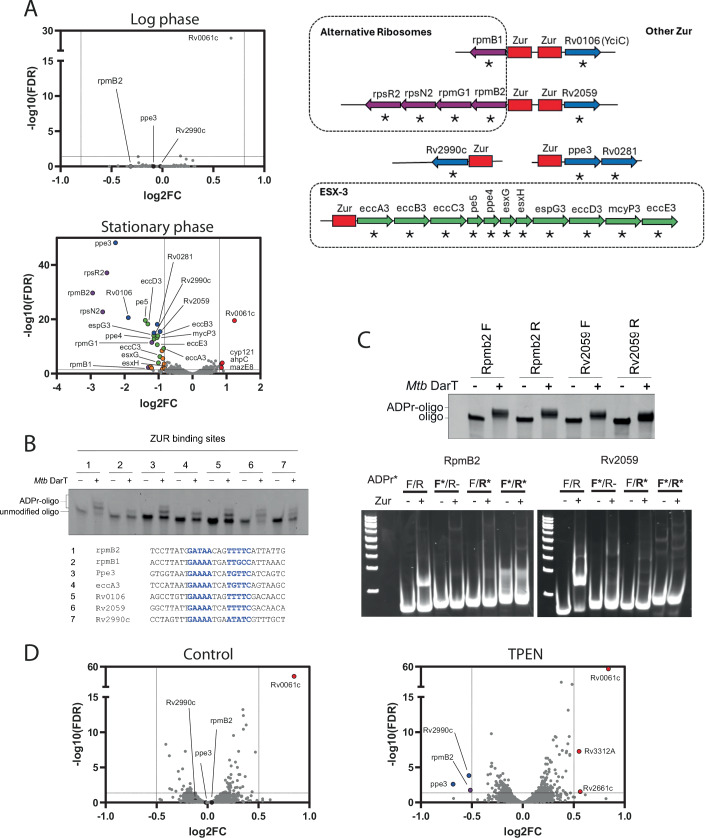
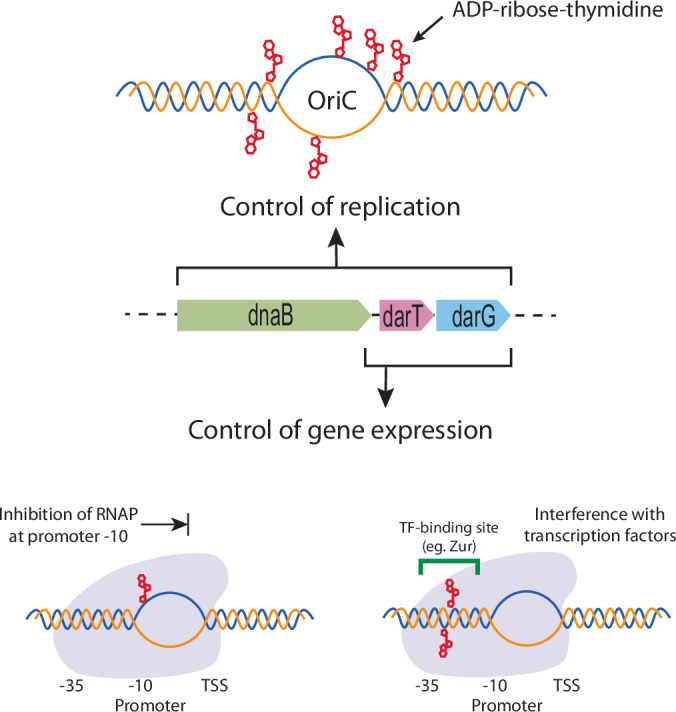
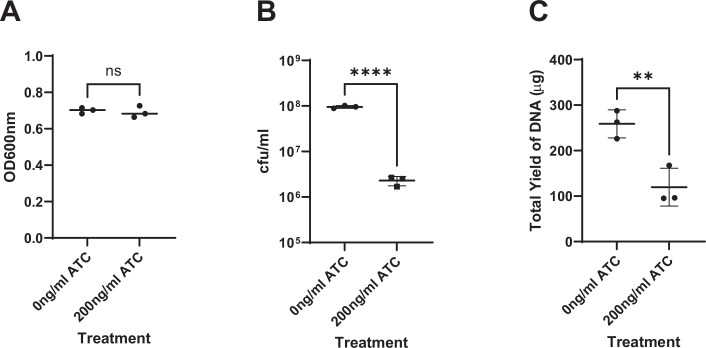
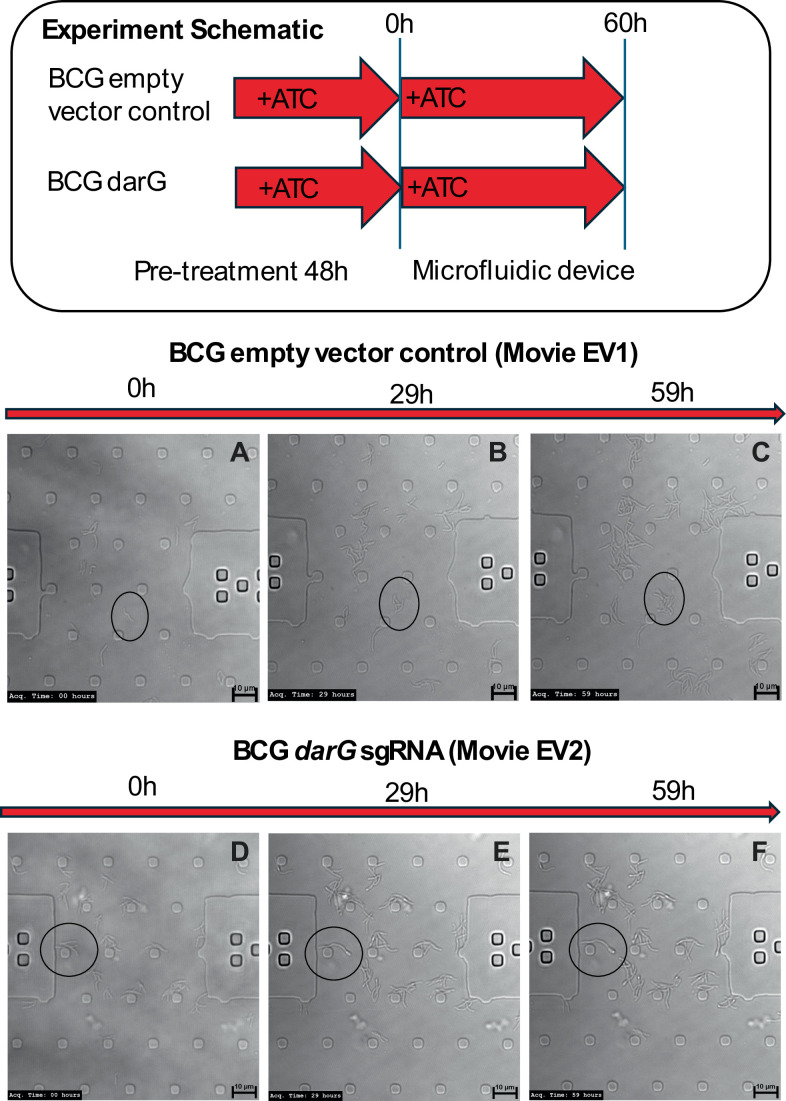
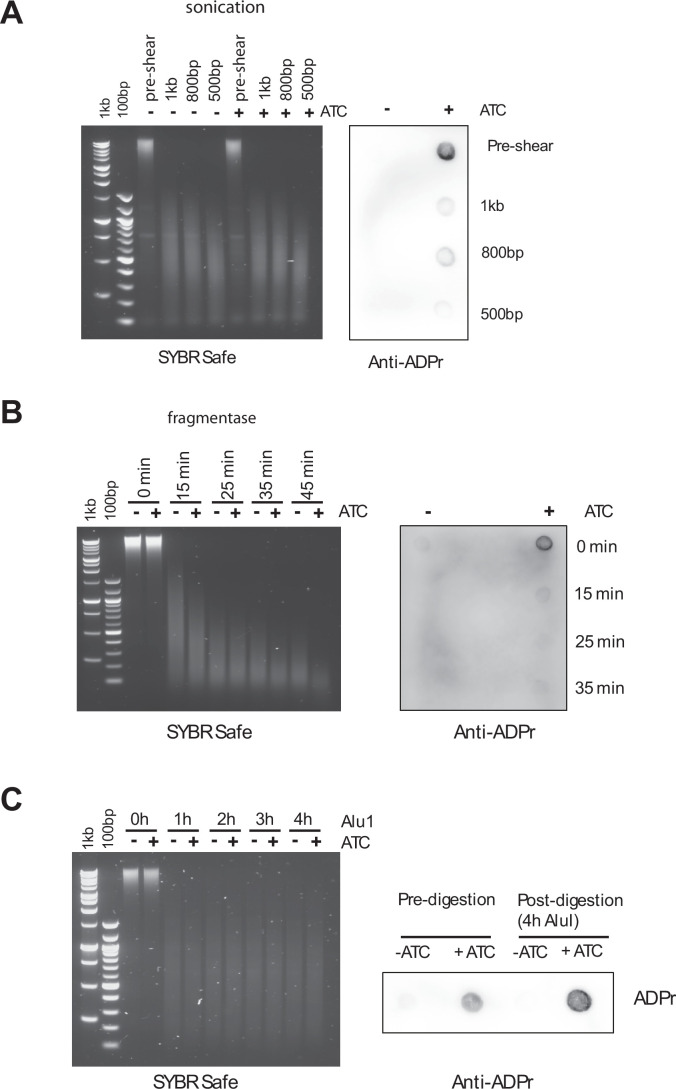
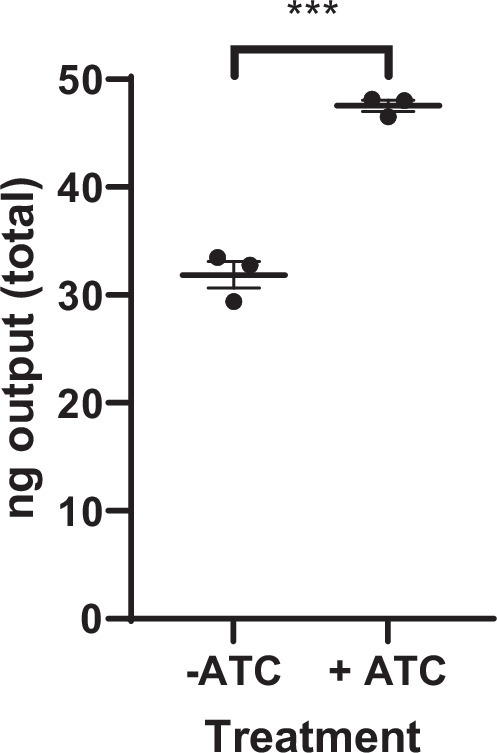
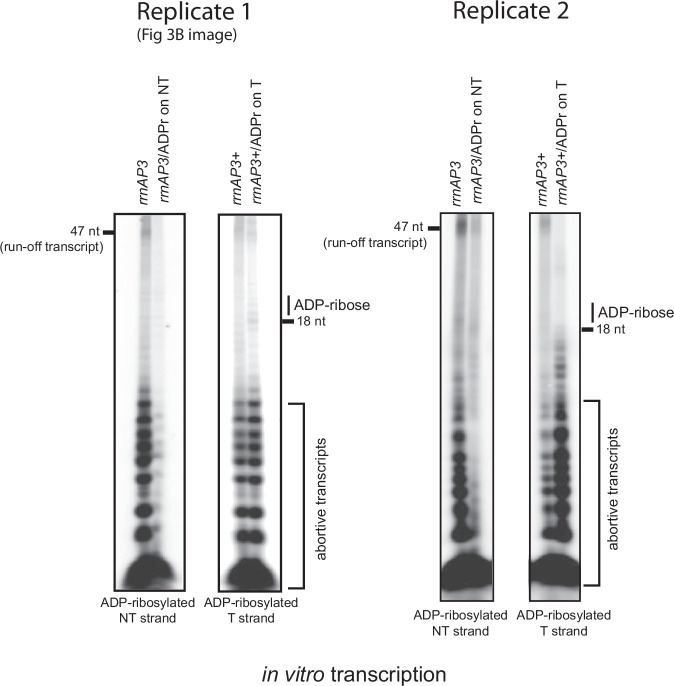
Mycobacterium tuberculosis maintains long-term infections characterised by the need to regulate growth and adapt to contrasting in vivo environments. Here we show that M. tuberculosis complex bacteria utilise reversible ADP-ribosylation of single-stranded DNA as a mechanism to coordinate stationary phase growth with transcriptional adaptation. The DNA modification is controlled by DarT, an ADP-ribosyltransferase, which adds ADP-ribose to thymidine, and DarG, which enzymatically removes this base modification. Using darG-knockdown M. bovis BCG, we map the first DNA ADP-ribosylome from any organism. We show that inhibition of replication by DarT is reversible and accompanied by extensive ADP-ribosylation at the origin of replication (OriC). In addition, we observe ADP-ribosylation across the genome and demonstrate that ADP-ribose-thymidine alters the transcriptional activity of M. tuberculosis RNA polymerase. Furthermore, we demonstrate that during stationary phase, DarT-dependent ADP-ribosylation of M. tuberculosis DNA is required to optimally induce expression of the Zur regulon, including the ESX-3 secretion system and multiple alternative ribosome proteins. Thus, ADP-ribosylation of DNA can provide a mechanistic link through every aspect of DNA biology from replication to transcription to translation.
Keywords: ADP-ribosylation; ADPr-Seq; DNA Modification; PARP; Transcription Regulation.
© 2025. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
