

Costs of reproduction in flowering plants
- PMID: 40342244
- PMCID: PMC12138173
- DOI: 10.1111/nph.70166
Costs of reproduction in flowering plants
Abstract
Costs of reproduction arise when investments into current reproduction reduce future reproductive fitness. Studies on reproductive costs use diverse approaches, including the analysis of gene expression, physiology, trade-offs between reproduction and growth/survival, and the impact of reproductive investments on population growth. These studies demonstrate that reproductive trade-offs have far-reaching effects on plants, affect their fitness, and are therefore important for shaping the evolution of life histories. However, not all studies have detected costs of reproduction, and c. 90% of these were conducted in natural populations, where controlling for variation in plant resource status is challenging. For dioecious plants, there is a common perception that fruit production should result in greater costs of reproduction for females than males, but divergent reproductive costs between the sexes are not supported by studies of reproductive trade-offs in dioecious plants. Other aspects of reproductive costs remain poorly understood, including ecological costs of reproduction, the fitness effects of reproductive trade-offs involving growth or physiological processes, and how the male sex role influences reproductive costs. Progress will be enabled by the use of measurements that allow for easier comparisons across studies and by more clearly distinguishing between the processes that contribute to current vs future reproductive fitness.
Keywords: cost function; demographic costs of reproduction; life‐history trade‐offs; reproductive costs; sex‐allocation theory; somatic costs of reproduction.
© 2025 The Author(s). New Phytologist © 2025 New Phytologist Foundation.
Conflict of interest statement
None declared.
Figures

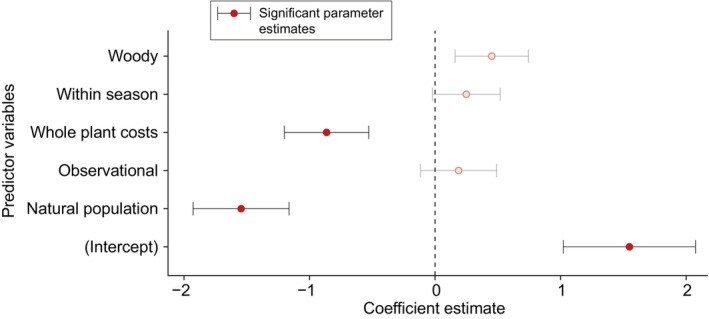

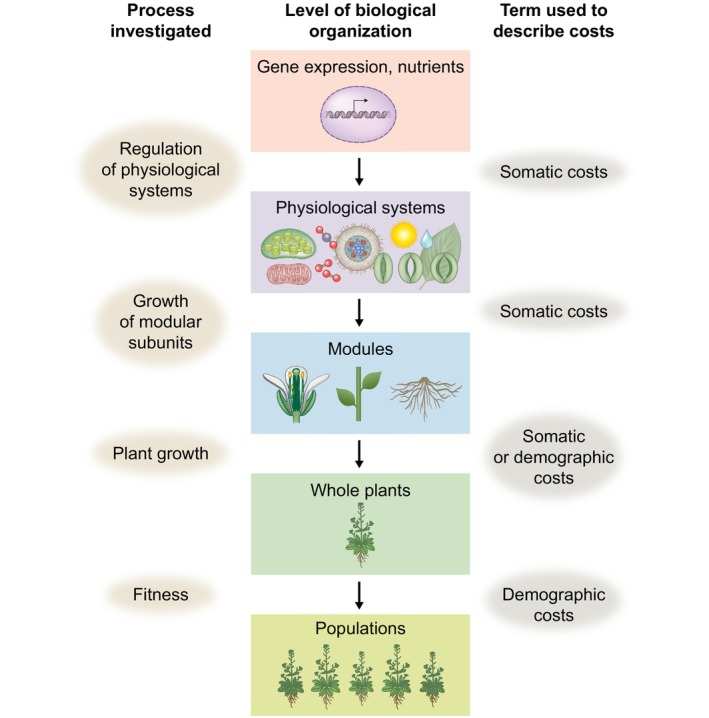
Similar articles
-
Gender-specific costs of reproduction on vegetative growth and physiological performance in the dioecious shrub Corema album.Ann Bot. 2010 Dec;106(6):989-98. doi: 10.1093/aob/mcq197. Epub 2010 Sep 29. Ann Bot. 2010. PMID: 20884627 Free PMC article.
-
Life-history trade-offs promote the evolution of dioecy.J Evol Biol. 2018 Sep;31(9):1405-1412. doi: 10.1111/jeb.13335. Epub 2018 Jul 4. J Evol Biol. 2018. PMID: 29908091
-
Reproductive allocation and costs in gynodioecious Leucopogon melaleucoides (Ericaceae): implications for the evolution of gender dimorphism.Plant Biol (Stuttg). 2011 Nov;13(6):888-95. doi: 10.1111/j.1438-8677.2011.00457.x. Epub 2011 Mar 4. Plant Biol (Stuttg). 2011. PMID: 21972998
-
The costs of reproduction in plants.New Phytol. 2002 Sep;155(3):321-348. doi: 10.1046/j.1469-8137.2002.00477.x. New Phytol. 2002. PMID: 33873312 Review.
-
The evolution of growth trajectories: what limits growth rate?Biol Rev Camb Philos Soc. 2011 Feb;86(1):97-116. doi: 10.1111/j.1469-185X.2010.00136.x. Biol Rev Camb Philos Soc. 2011. PMID: 20394607 Review.
References
-
- Acerenza L. 2016. Constraints, trade‐offs and the currency of fitness. Journal of Molecular Evolution 82: 117–127. - PubMed
-
- Ågren J, Willson M. 1994. Cost of seed production in the perennial herbs Geranium maculatum and G. sylvaticum: an experimental field study. Oikos 70: 35–42.
-
- Aragón CF, Méndez M, Escudero A. 2009. Survival costs of reproduction in a short‐lived perennial plant: live hard, die young. American Journal of Botany 96: 904–911. - PubMed
-
- Ashman T‐L. 1994. A dynamic perspective on the physiological cost of reproduction in plants. American Naturalist 144: 300–316.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
