

A retinoic acid:YAP1 signaling axis controls atrial lineage commitment
- PMID: 40343798
- PMCID: PMC12186250
- DOI: 10.1016/j.celrep.2025.115687
A retinoic acid:YAP1 signaling axis controls atrial lineage commitment
Abstract
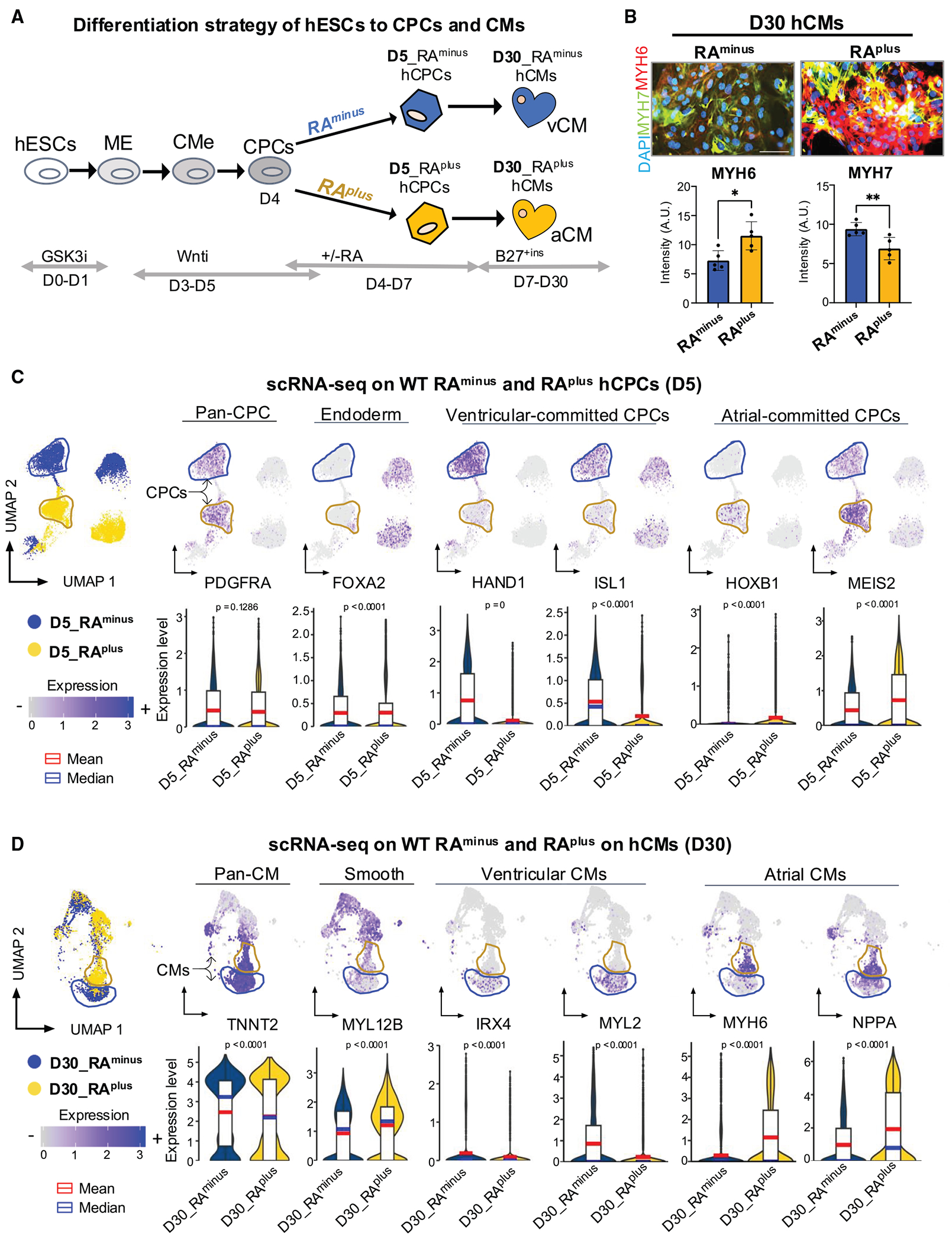
In cardiac progenitor cells (CPCs), retinoic acid (RA) signaling induces atrial lineage gene expression and acquisition of an atrial cell fate. To achieve this, RA coordinates a complex regulatory network of downstream effectors that is not fully identified. To address this gap, we applied a functional genomics approach (i.e., scRNA-seq and snATAC-seq) to untreated and RA-treated human embryonic stem cell (hESC)-derived CPCs. Unbiased analysis revealed that the Hippo effectors YAP1 and TEAD4 are integrated with the atrial transcription factor enhancer network and that YAP1 activates RA enhancers in CPCs. Furthermore, Yap1 deletion in mouse embryos compromises the expression of RA-induced genes, such as Nr2f2, in the CPCs of the second heart field. Accordingly, in hESC-derived patterned heart organoids, YAP1 regulates the formation of an atrial chamber but is dispensable for the formation of a ventricle. Overall, our findings revealed that YAP1 cooperates with RA signaling to induce atrial lineages during cardiogenesis.
Keywords: CP: Developmental biology; NR2F2; SHF; TEAD4; YAP1; atrial differentiation; cardiac enhancers; cardiac progenitors; chamber development; heart organoids; retinoic acid signaling.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Update of
-
A Retinoic Acid:YAP1 signaling axis controls atrial lineage commitment.bioRxiv [Preprint]. 2024 Jul 12:2024.07.11.602981. doi: 10.1101/2024.07.11.602981. bioRxiv. 2024. Update in: Cell Rep. 2025 May 27;44(5):115687. doi: 10.1016/j.celrep.2025.115687. PMID: 39026825 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
