

Mitochondria regulate MR1 protein expression and produce self-metabolites that activate MR1-restricted T cells
- PMID: 40354545
- PMCID: PMC12107159
- DOI: 10.1073/pnas.2418525122
Mitochondria regulate MR1 protein expression and produce self-metabolites that activate MR1-restricted T cells
Abstract
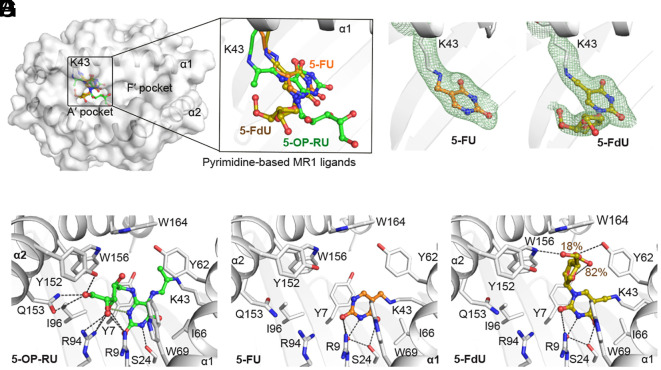
Mitochondria coordinate several metabolic pathways, producing metabolites that influence the immune response in various ways. It remains unclear whether mitochondria impact antigen presentation by the MHC-class-I-related antigen-presenting molecule, MR1, which presents small molecules to MR1-restricted T-lymphocytes. Here, we demonstrate that mitochondrial complex III and the enzyme dihydroorotate dehydrogenase are essential for the cell-surface expression of MR1 and for generating uridine- and thymidine-related compounds that bind to MR1 and are produced upon oxidation by reactive oxygen species. One mitochondria-derived immunogenic formylated metabolite we identified is 5-formyl-deoxyuridine (5-FdU). Structural studies indicate that 5-FdU binds in the A'-antigen-binding pocket of MR1, positioning the deoxyribose toward the surface of MR1 for TCR interaction. 5-FdU stimulates specific T cells and detects circulating T cells when loaded onto MR1-tetramers. 5-FdU-reactive cells resemble adaptive T cells and express the phenotypes of naïve, memory, and effector cells, indicating prior in vivo stimulation. These findings suggest that mitochondria may play a role in MR1-mediated immune surveillance.
Keywords: MR1; T cells; antigen presentation; formylated metabolite; mitochondria.
Conflict of interest statement
Competing interests statement:A.V., A.C., L.M., and G.D.L. are inventors on patent applications submitted by the University of Basel covering the use of MR1T cells and nucleobase adducts for immunomodulation. J.R. is an inventor on patents describing MR1 ligands and MR1 tetramers. All other authors declare no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
- DE220101491/Australian Cancer Research Foundation (ACRF)
- 310030-173240/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (SNF)
- KLbB-4779-02-2019/Krebsliga Beider Basel (Cancer League of Basel-City and Basel-Country)
- 310030B-192828/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (SNF)
- RO1 AI148407-01A1/HHS | NIH (NIH)
LinkOut - more resources
Full Text Sources
Research Materials
