

Gasdermin-D pores induce an inactivating caspase-4 cleavage that limits IL-18 production in the intestinal epithelium
- PMID: 40355718
- PMCID: PMC12069520
- DOI: 10.1038/s42003-025-08183-9
Gasdermin-D pores induce an inactivating caspase-4 cleavage that limits IL-18 production in the intestinal epithelium
Erratum in
-
Author Correction: Gasdermin-D pores induce an inactivating caspase-4 cleavage that limits IL-18 production in the intestinal epithelium.Commun Biol. 2025 May 31;8(1):842. doi: 10.1038/s42003-025-08266-7. Commun Biol. 2025. PMID: 40450095 Free PMC article. No abstract available.
Abstract
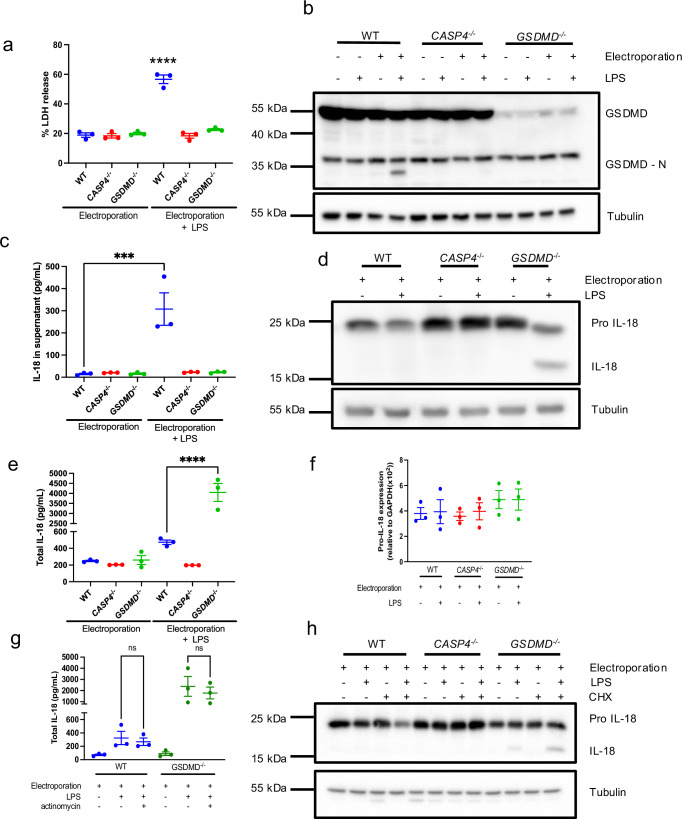
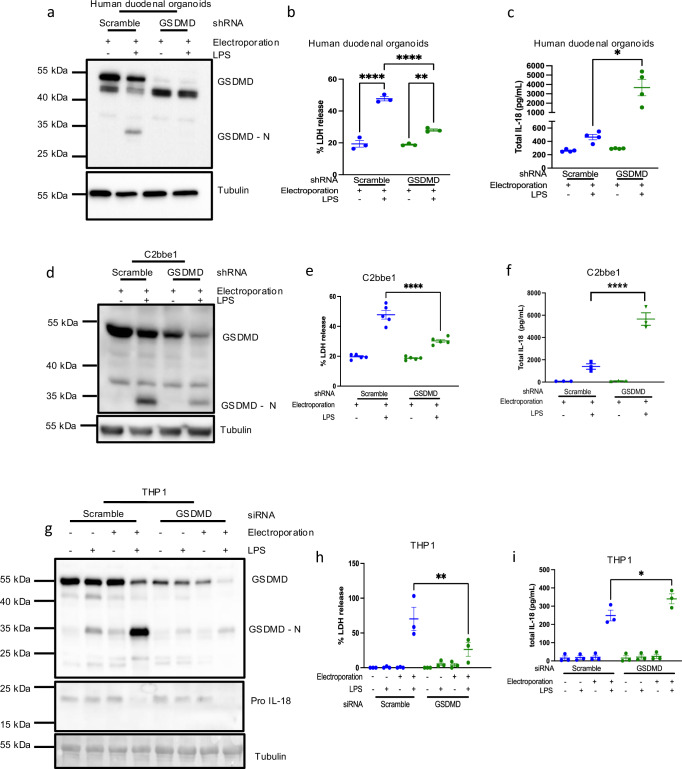
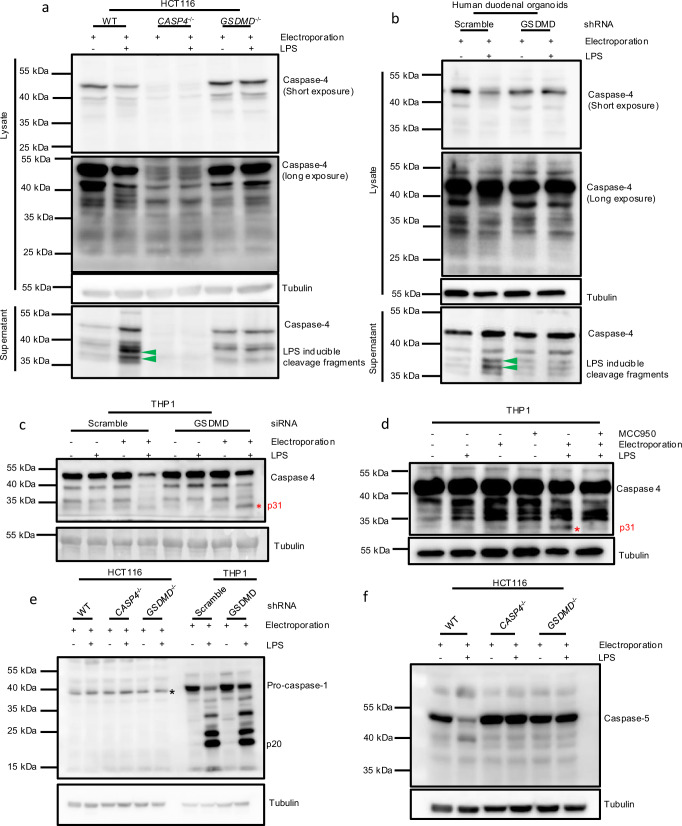
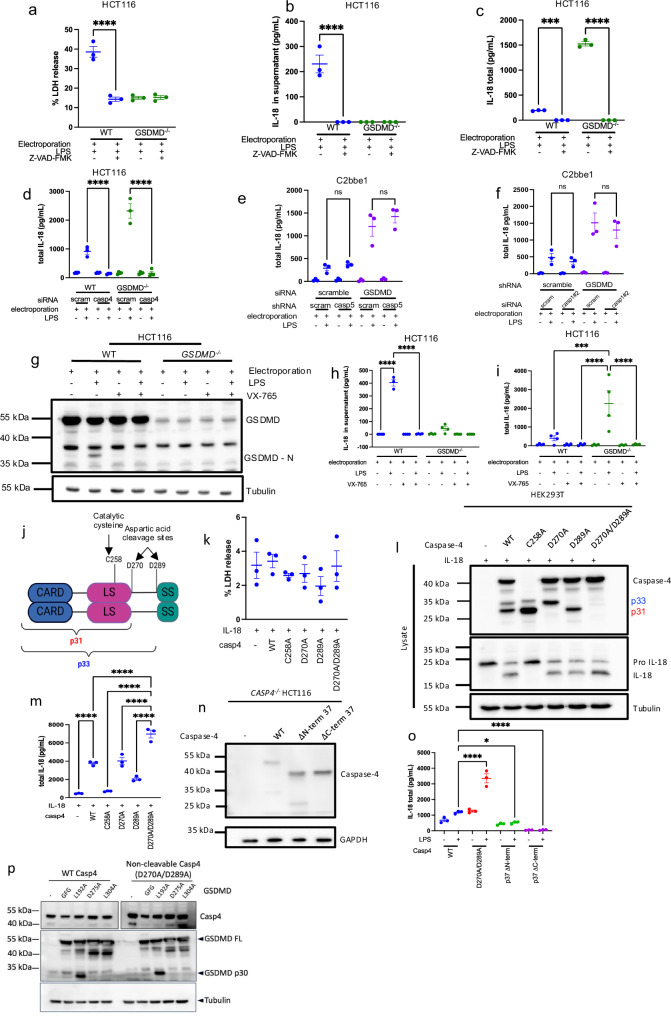
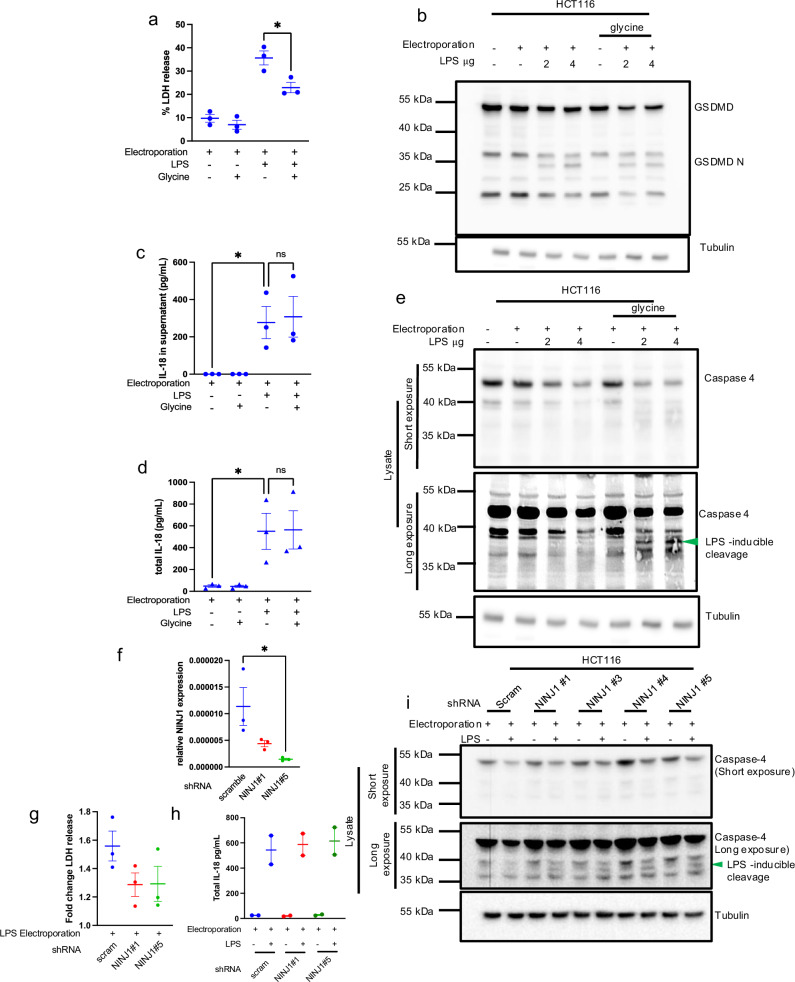
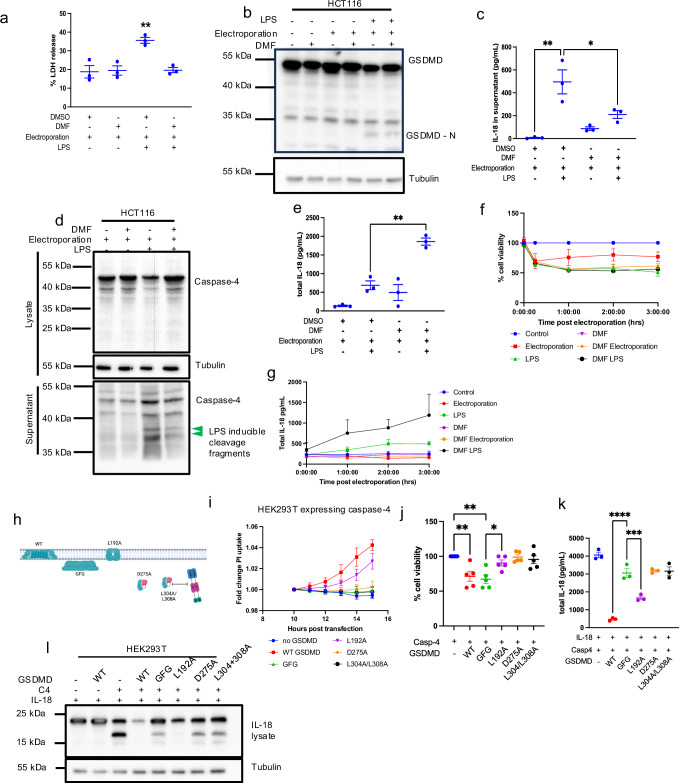
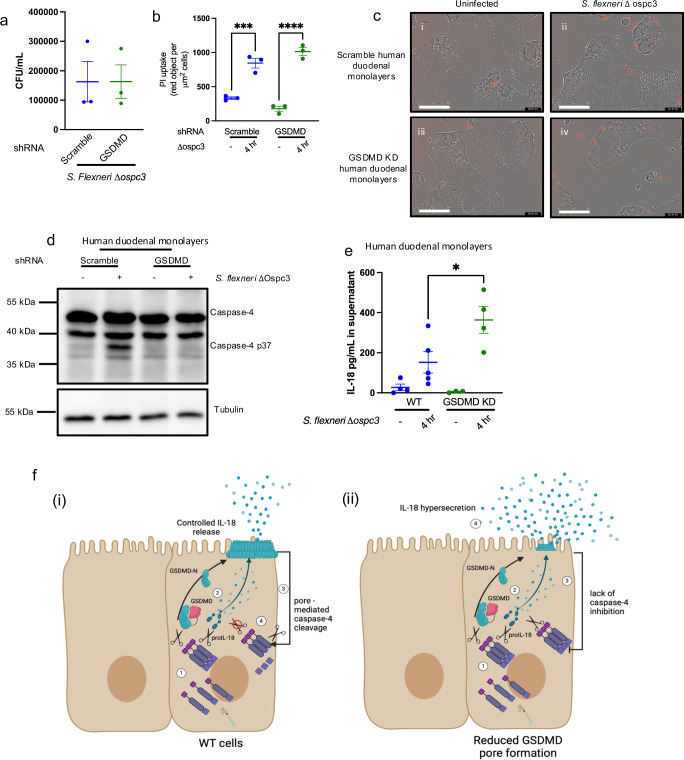
Intestinal epithelial-derived IL-18 is critical for homeostatic intestinal barrier function and is secreted through Gasdermin D (GSDMD) pores. Inflammasome activation is a prerequisite for both IL-18 maturation and GSDMD pore formation. However, GSDMD pores also cause pyroptotic cell death, which could be detrimental to the intestinal epithelial barrier. How epithelial cells balance the need to secrete IL-18 and to maintain barrier integrity remains poorly understood. In human intestinal epithelial cell lines and in primary human epithelial intestinal organoids, but not in immune cells, GSDMD plasma membrane pore formation by LPS electroporation and by gram-negative bacterial infection induced a non-conventional p37 caspase-4 fragment that was associated with reduced levels of mature IL-18. By contrast, limiting GSDMD plasma membrane pores pharmacologically and via point-mutagenesis prevented caspase-4 cleavage and increased IL-18 production, suggesting that p37 caspase-4 cleavage may regulate IL-18 maturation in the intestinal epithelium. In support, co-expression of caspase-4 cleavage mutants and IL-18 in HEK293T cells revealed that non-cleavable caspase-4 produced more mature IL-18 than cleaved caspase-4. Overall, these studies suggest that epithelial inflammasomes encode feedback pathways that control the balance between cytokine secretion and cell death. This may be an important mechanism to ensure homeostatic IL-18 production in the intestinal epithelium.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
-
- Winsor, N., Krustev, C., Bruce, J., Philpott, D. J. & Girardin, S. E. Canonical and noncanonical inflammasomes in intestinal epithelial cells. Cell Microbiol21, e13079 (2019). - PubMed
-
- Shi, J. et al. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature526, 660–665 (2015). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
