

Melatonin enhances everolimus efficacy in breast cancer by suppressing mTOR pathway activation and promoting apoptosis and mitochondrial function
- PMID: 40355936
- PMCID: PMC12070795
- DOI: 10.1186/s40360-025-00907-1
Melatonin enhances everolimus efficacy in breast cancer by suppressing mTOR pathway activation and promoting apoptosis and mitochondrial function
Abstract
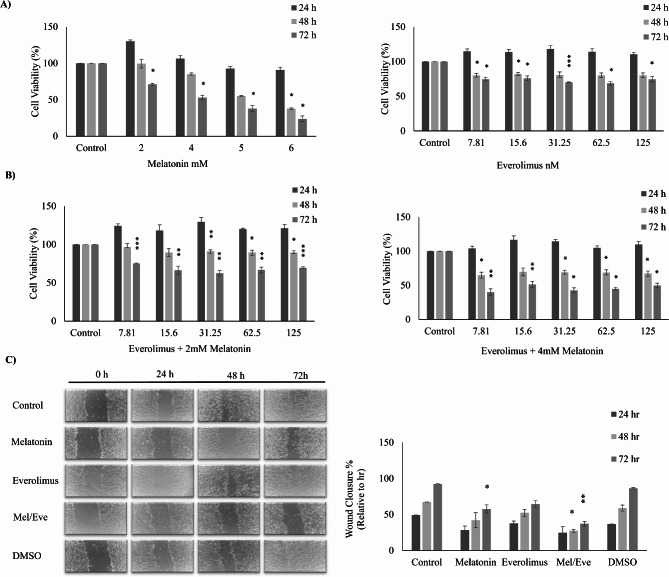
Background: Everolimus is used in the treatment of breast cancer by targeting the PI3K/AKT/mTOR pathway, particularly during anti-hormonal therapy. The efficacy of everolimus is limited due to a feedback loop that supresses mTOR while simultaneously enhancing Akt activation in endocrine-resistant breast cancer. Melatonin (N-acetyl-5-methoxytryptamine) regulates mitochondrial activity, cell death, and autophagy due to its strong free radical scavenging, antioxidant, and anti-inflammatory characteristics. Melatonin, a naturally occurring oncostatic agent, slows tumor growth in a range of malignancies, including breast cancer. Due to its ability to protect healthy cells from oxidative stress and inflammation, along with its anti-cancer properties, melatonin has the potential to serve asan effective adjuvant in breast cancer therapy. It also inhibits the phosphorylation of mTOR and Akt, two essential pathways implicated in breast cancer growth, which may aid in overcoming resistance to targeted treatments like everolimus. The combination effects of melatonin and everolimus on hormone receptor-positive breast cancer remains unexplored. This study examined the effectiveness of melatonin when combined with everolimus for the treatment of hormone receptor-positive breast cancer.
Methods: To investigate the effects of melatonin and everolimus combination, we divided MCF-7 cells into four experimental groups: the control, Melatonin (3 mM), Everolimus (30 nM), and a combination of Melatonin and Everolimus (3 mM + 30 nM). Cell viability, apoptosis, autophagy activation, and mitochondrial function were evaluated using established techniques.
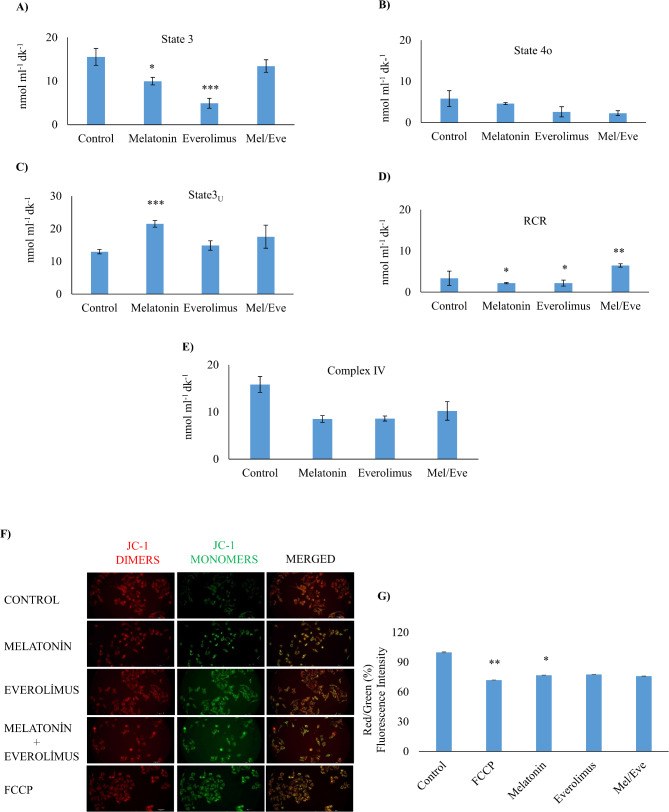
Results: Based on the cell viability test, the combination of 30 nM everolimus and 3 mM melatonin inhibited phosphorylation of 4E-BP1 and p70S6K, which are downstream effectors of the mTOR pathway, and reduced cell growth. In addition, co-administration of melatonin and everolimus increased apoptosis and led to Sub-G1 phase accumulation. LC3 protein expression and LC3 puncta analysis demonstrated autophagic activity. In terms of mitochondrial function, co-administration of melatonin with everolimus did not cause proton leakage or mitochondrial uncoupling, but did restore everolimus-induced respiratory inhibition.
Conclusions: In conclusion, melatonin is thought to improve the effectiveness of everolimus by inhibiting mTOR downstream effectors, enhancing apoptosis, activating autophagy, improving mitochondrial respiration, and reducing MCF-7 growth.
Keywords: Apoptosis; Autophagy; Everolimus; Melatonin; Mitochondrial respiration; mTOR.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval: The author of this article declares that the materials and methods used in this study do not require ethical committee permission and/or legal-special permissions. Consent to participate: Not applicable. Consent for publication: All authors give their consent for the publication of the data/manuscript. Competing interests: The authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
