

Aneuploidy enables adaptation to brefeldin A in Candida albicans
- PMID: 40357392
- PMCID: PMC12066683
- DOI: 10.3389/fcimb.2025.1562726
Aneuploidy enables adaptation to brefeldin A in Candida albicans
Abstract
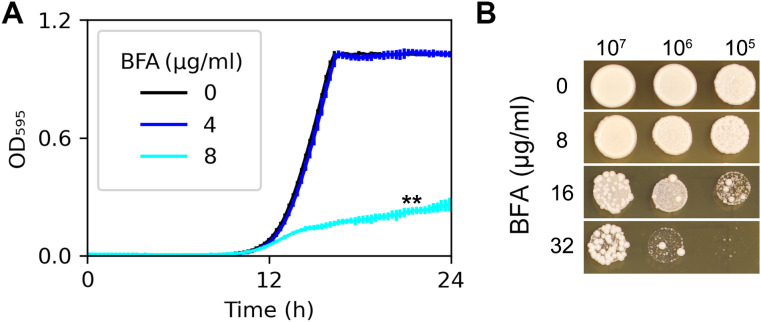
Candida albicans is the most prevalent opportunistic fungal pathogen. Both in vivo and in vitro studies have demonstrated that genome plasticity is a hallmark of C. albicans. While aneuploidy formation is a well-documented adaptive mechanism under various stress conditions, the response to brefeldin A-a compound that induces endoplasmic reticulum stress-remains poorly understood. In this study, we demonstrate that C. albicans adapts to subinhibitory and inhibitory concentrations of brefeldin A, primarily through the formation of chromosome 3 trisomy. These aneuploid strains were found to be unstable, reverting to euploidy in the absence of stress, accompanied by a loss of brefeldin A tolerance. We identified at least two genes on chromosome 3, SEC7 and CDR1, that contribute to this adaptive response. Notably, higher concentrations of brefeldin A selected for strains with increasingly complex aneuploidies. Our findings underscore the remarkable genomic plasticity of C. albicans and reveal aneuploidy as a reversible mechanism for adapting to brefeldin A stress. This study provides new insights into the role of aneuploidy in fungal adaptation and offers potential implications for understanding drug resistance mechanisms in pathogenic fungi.
Keywords: Candida albicans; ER stress; aneuploidy; brefeldin A; genome instability.
Copyright © 2025 Wang, Wang, Dong, Yang and Xu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources