

DNGR-1 regulates proliferation and migration of bone marrow dendritic cell progenitors
- PMID: 40358588
- PMCID: PMC12071193
- DOI: 10.1084/jem.20241813
DNGR-1 regulates proliferation and migration of bone marrow dendritic cell progenitors
Abstract
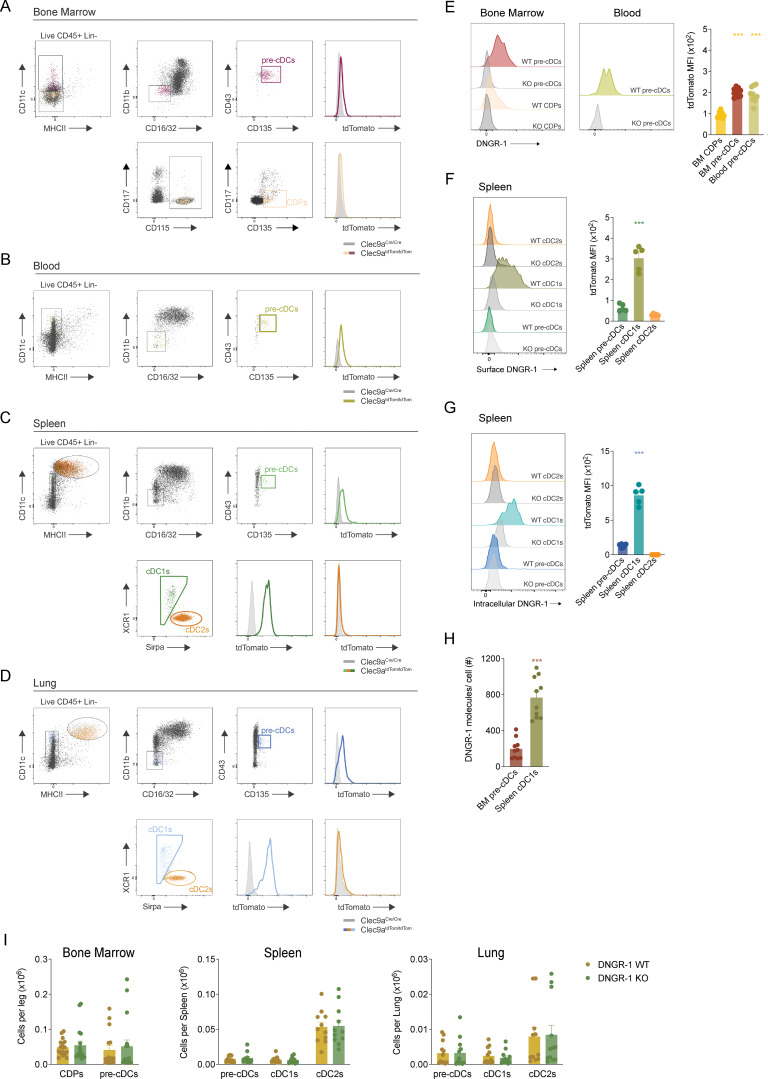
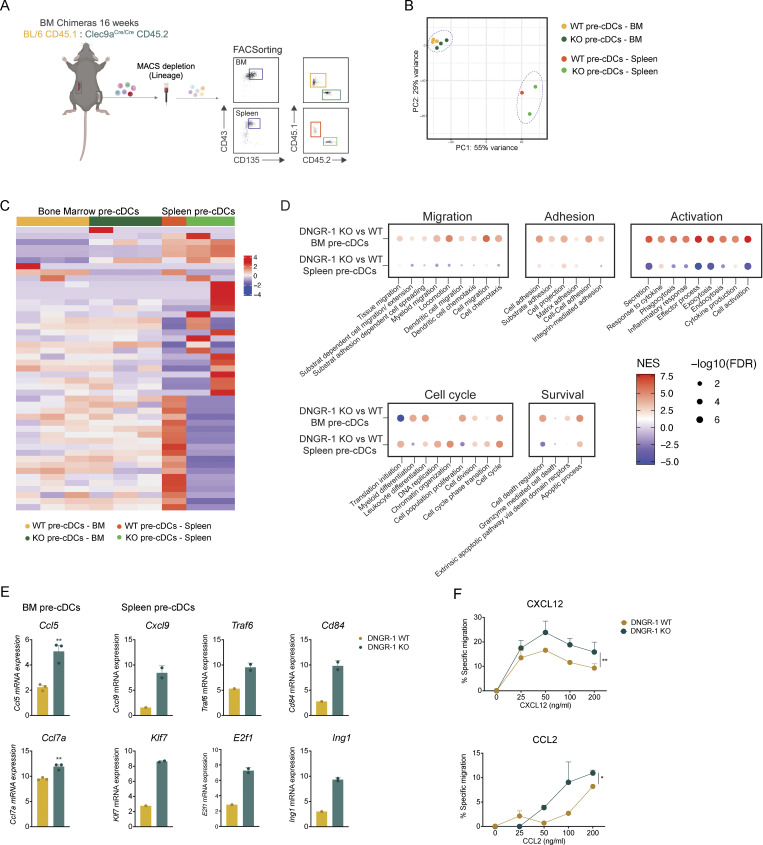
Conventional dendritic cells (cDCs) are sentinel cells that play a crucial role in both innate and adaptive immune responses. cDCs originate from a progenitor (pre-cDC) in the bone marrow (BM) that travels via the blood to seed peripheral tissues before locally differentiating into functional cDC1 and cDC2 cells, as part of a process known as cDCpoiesis. How cDCpoiesis is regulated and whether this affects the output of cDCs is poorly understood. In this study, we show that DNGR-1, an innate immune receptor expressed by cDC progenitors and type 1 cDCs, can regulate cDCpoiesis in mice. In a competitive chimera setting, cDC progenitors lacking DNGR-1 exhibit increased proliferation and tissue migratory potential. Compared with their WT counterparts, DNGR-1-deficient cDC progenitor cells display superior colonization of peripheral tissues but an altered distribution. These findings suggest that cDCpoiesis can be regulated in part by precursor cell-intrinsic processes driven by signals from innate immune receptors such as DNGR-1 that may respond to alterations in the BM milieu.
© 2025 Cardoso et al.
Conflict of interest statement
Disclosures: K.H.J. Lim reported funding by Wellcome Imperial 4i Clinical Research Fellowship and the Crick Clinical Postdoctoral Fellowship. C. Reis e Sousa reported personal fees from Adendra Therapeutics, Montis Biosciences, and Bicycle Therapeutics outside the submitted work; and additional appointments as Visiting Professor in the Faculty of Medicine at Imperial College London and at King’s College London and an honorary professorship at University College London. No other disclosures were reported.
Figures





References
-
- Ahrens, S., Zelenay S., Sancho D., Hanč P., Kjær S., Feest C., Fletcher G., Durkin C., Postigo A., Skehel M., et al. 2012. F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells. Immunity. 36:635–645. 10.1016/j.immuni.2012.03.008 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
