

Staphylococcus aureus SaeR/S-regulated factors overcome human complement-mediated inhibition of aggregation to evade neutrophil killing
- PMID: 40359050
- PMCID: PMC12107147
- DOI: 10.1073/pnas.2412447122
Staphylococcus aureus SaeR/S-regulated factors overcome human complement-mediated inhibition of aggregation to evade neutrophil killing
Abstract
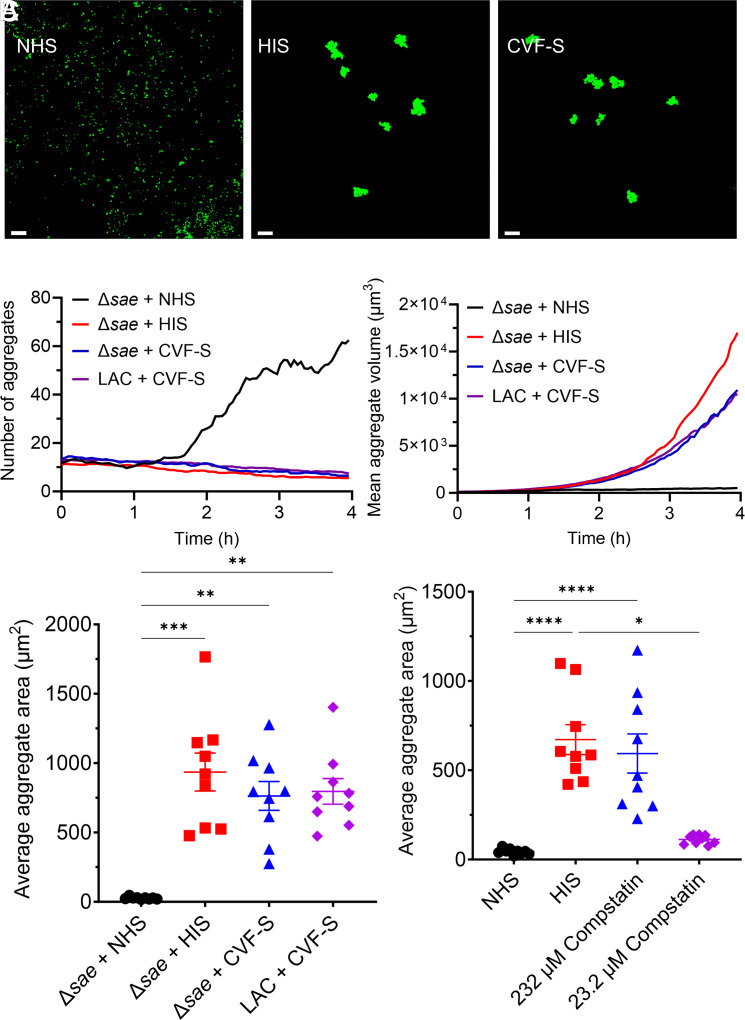
Staphylococcus aureus (S. aureus) is a frequent culprit in implant-associated infections and employs many virulence factors to escape killing by the host immune system. The specific immune evasion strategies used by small aggregates of S. aureus on a surface, precursors to mature biofilm, are still relatively unknown. Time-lapse confocal microscopy was leveraged to quantify interactions between S. aureus aggregates and human neutrophils in vitro and identify specific mechanisms of resistance to neutrophil killing. Surface-associated wild-type S. aureus rapidly formed small biofilm aggregates when grown in human serum. Conversely, aggregation was inhibited when the SaeR/S two-component gene regulatory system was deleted. Wild-type aggregates began to show individual and population-level resistance to neutrophil killing upon reaching sizes of approximately 50 to 75 µm2, whereas Δsae clusters failed to reach these sizes and were readily cleared. Aggregation of Δsae strains was impaired by serum complement, and this inhibition required complement proteins C3 and factor B, but not C4 or C5, suggesting that this activity primarily occurs at the level of the alternative pathway. Several complement-inhibiting genes regulated by SaeR/S were identified that collectively facilitate biofilm aggregate formation in human, but not murine serum. Finally, aggregation of two related opportunistic pathogens, Staphylococcus epidermidis and Enterococcus faecalis, was inhibited by serum. These data demonstrate a function of serum complement, the ability to inhibit bacterial aggregation, that is potently blocked by S. aureus through the production of multiple complement-interfering proteins that are regulated by the SaeR/S system.
Keywords: Staphylococcus aureus; biofilm; complement; microscopy; neutrophil.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- Costerton J. W., Stewart P. S., Greenberg E. P., Bacterial biofilms: A common cause of persistent infections. Science 284, 1318–1322 (1999). - PubMed
-
- Parsek M. R., Singh P. K., Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 57, 677–701 (2003). - PubMed
-
- Arciola C. R., An Y. H., Campoccia D., Donati M. E., Montanaro L., Etiology of implant orthopedic infections: A survey on 1027 clinical isolates. Int. J. Artif. Organs 28, 1091–1100 (2005). - PubMed
-
- Arciola C. R., Campoccia D., Montanaro L., Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 16, 397–409 (2018). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
