

Control of spatiotemporal activation of organ-specific fibers in the swine vagus nerve by intermittent interferential current stimulation
- PMID: 40360488
- PMCID: PMC12075724
- DOI: 10.1038/s41467-025-59595-4
Control of spatiotemporal activation of organ-specific fibers in the swine vagus nerve by intermittent interferential current stimulation
Abstract
Vagus nerve stimulation (VNS) is emerging as potential treatment for several chronic diseases. However, limited control of fiber activation, e.g., to promote desired effects over side effects, restricts clinical translation. Towards that goal, we describe a VNS method consisting of intermittent, interferential sinusoidal current stimulation (i2CS) through multi-contact epineural cuffs. In experiments in anesthetized swine, i2CS elicits nerve potentials and organ responses, from lungs and laryngeal muscles, that are distinct from equivalent non-interferential sinusoidal stimulation. Resection and micro-CT imaging of a previously stimulated nerve, to resolve anatomical trajectories of nerve fascicles, demonstrate that i2CS responses are explained by activation of organ-specific fascicles rather than the entire nerve. Physiological responses in swine and activity of single fibers in anatomically realistic, physiologically validated biophysical vagus nerve models indicate that i2CS reduces fiber activation at the interference focus. Experimental and modeling results demonstrate that current steering and beat and repetition frequencies predictably shape the spatiotemporal pattern of fiber activation, allowing tunable and precise control of nerve and organ responses. When compared to equivalent sinusoidal stimulation in the same animals, i2CS produces reduced levels of a side-effect by larger laryngeal fibers, while attaining similar levels of a desired effect by smaller bronchopulmonary fibers.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: IMEC has been granted patents related to this work (US18511762, US0017074A1, US0364428A1, US0198109A1). IMEC and Northwell have submitted patents related to this work.
Figures
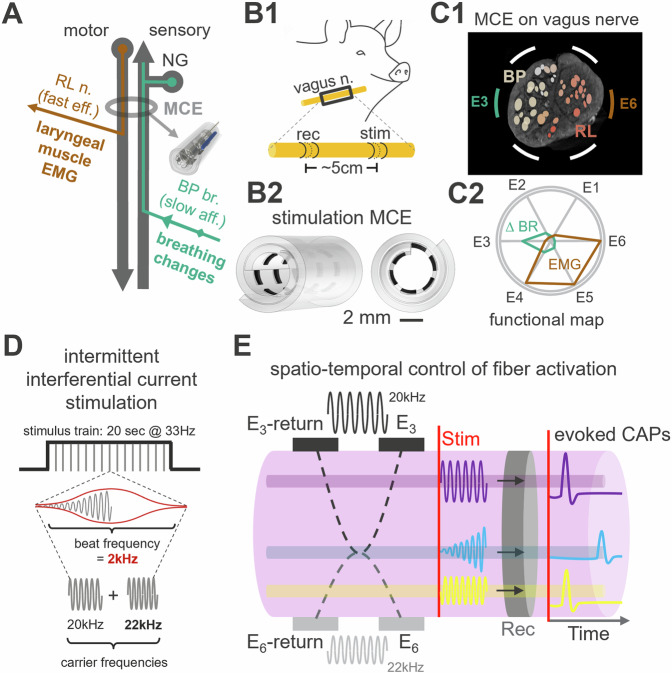

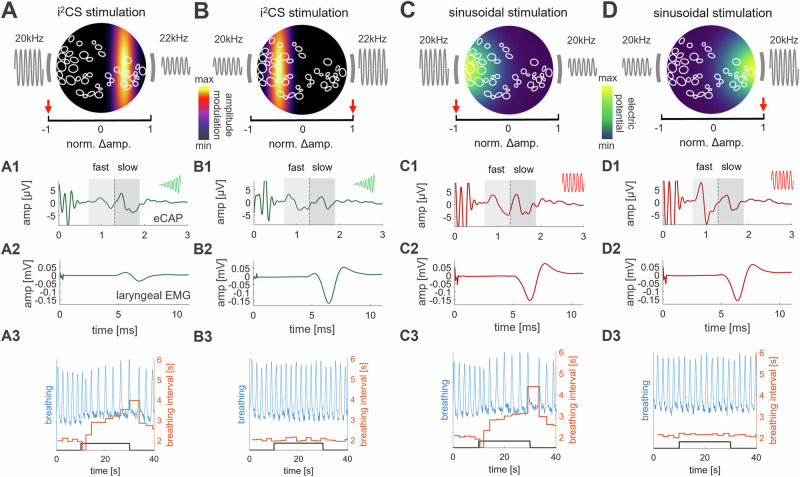




References
-
- Jänig, W. The Integrative Action of the Autonomic Nervous System: Neurobiology of Homeostasis. 10.1017/9781108778411 (Cambridge University Press, Cambridge, 2022).
-
- Karemaker, J. M. The multibranched nerve: vagal function beyond heart rate variability. Biol. Psychol.172, 108378 (2022). - PubMed
-
- Ben-Menachem, E. Vagus nerve stimulation, side effects, and long-term safety. J. Clin. Neurophysiol.18, 415–418 (2001). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
