

Epstein-Barr virus latent membrane protein 1 subverts IMPDH pathways to drive B-cell oncometabolism
- PMID: 40367275
- PMCID: PMC12169587
- DOI: 10.1371/journal.ppat.1013092
Epstein-Barr virus latent membrane protein 1 subverts IMPDH pathways to drive B-cell oncometabolism
Abstract
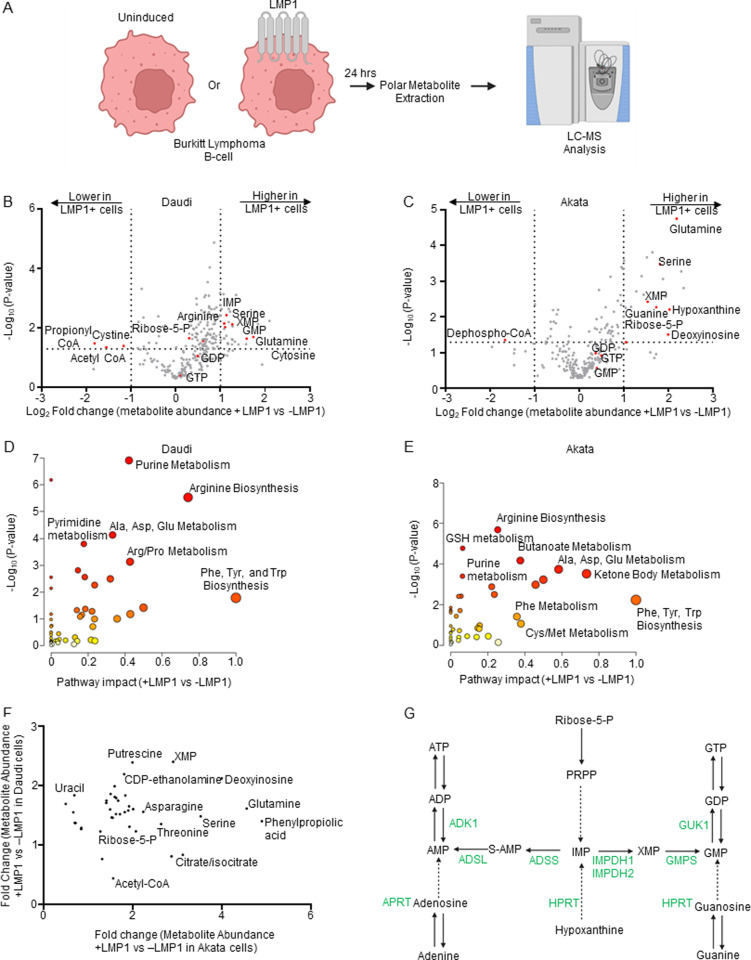
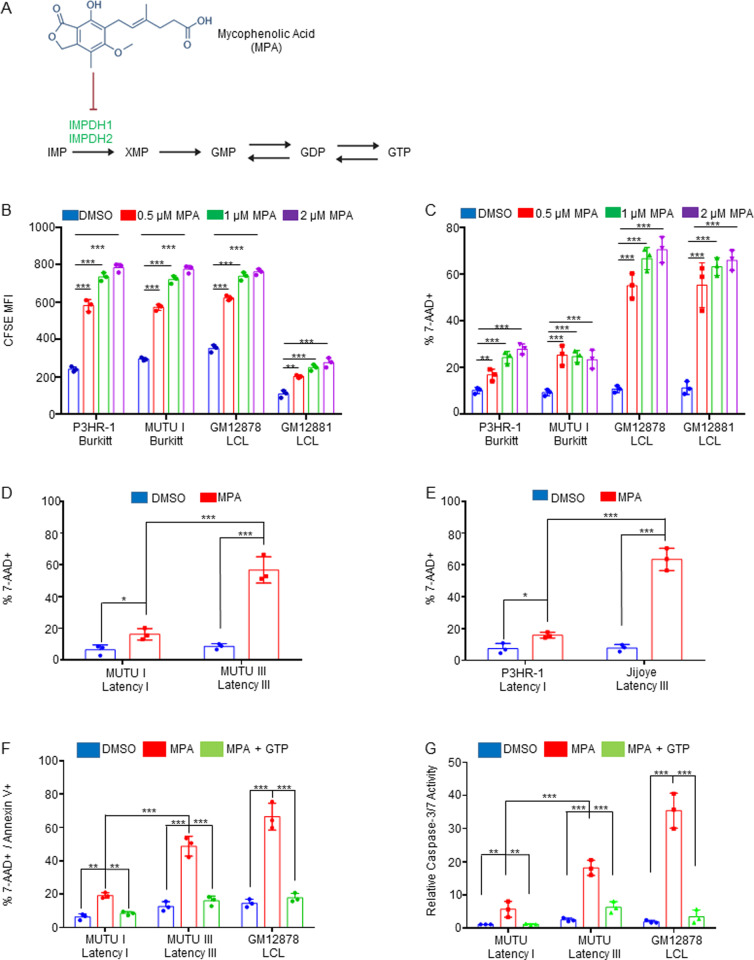
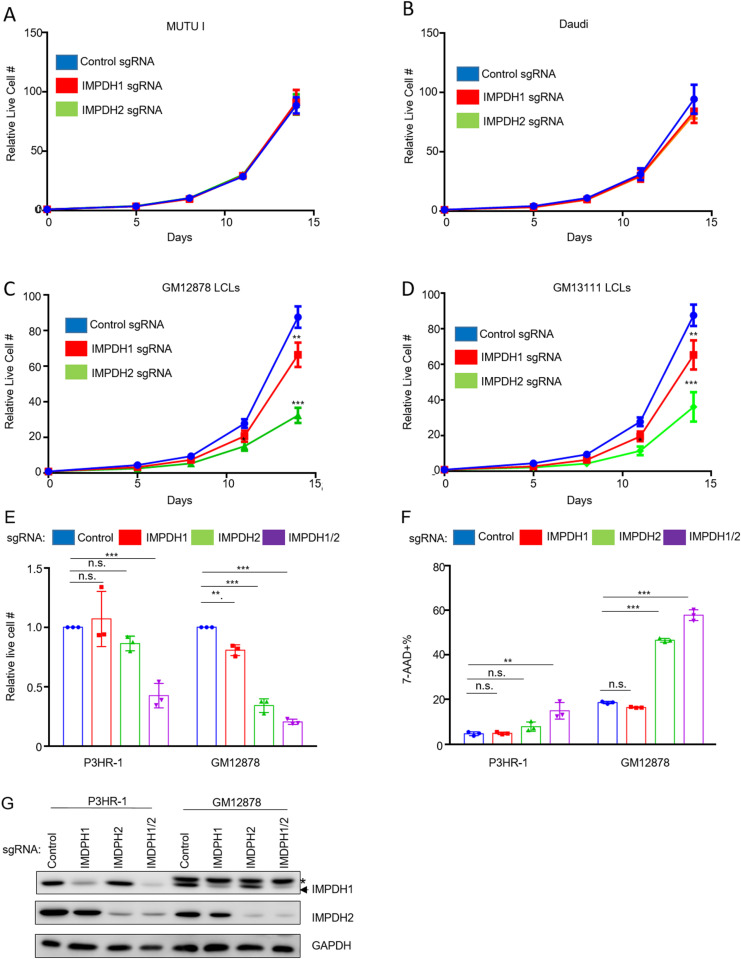
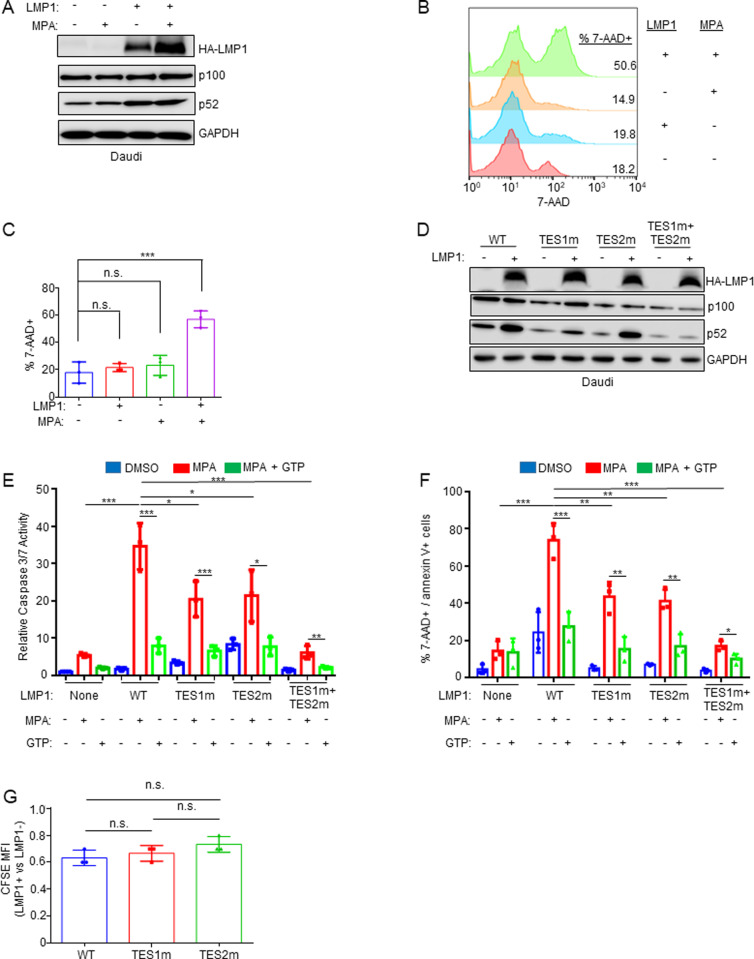
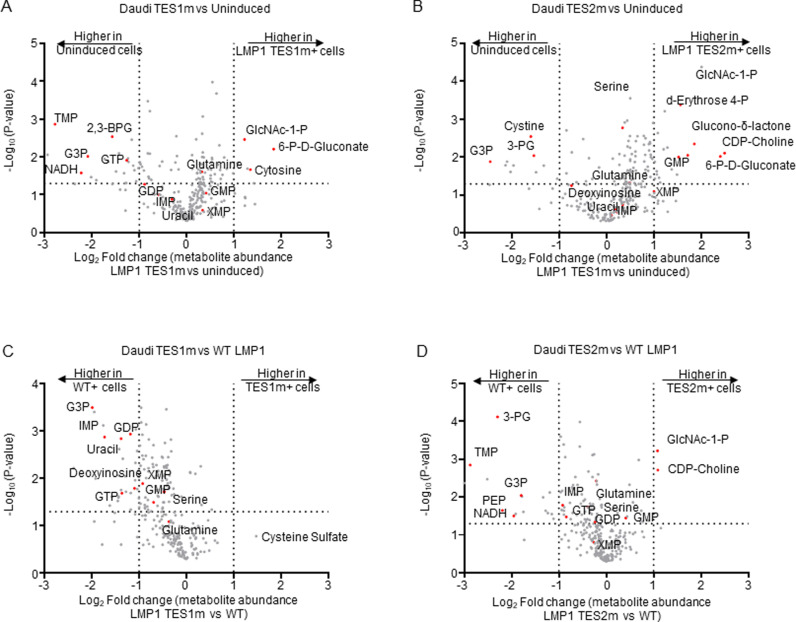
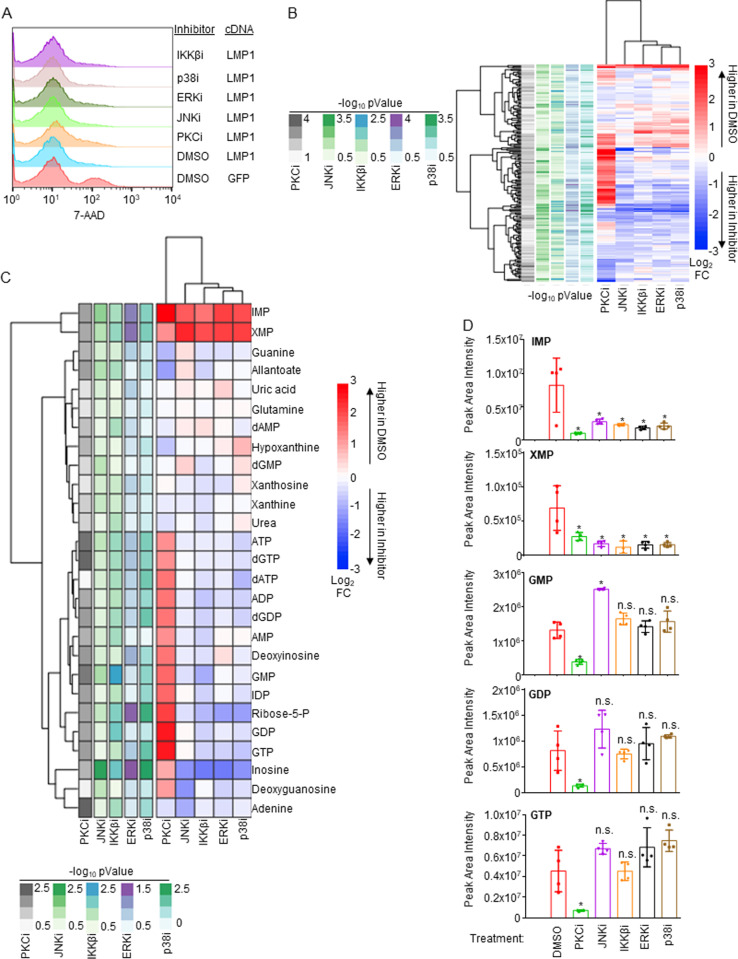
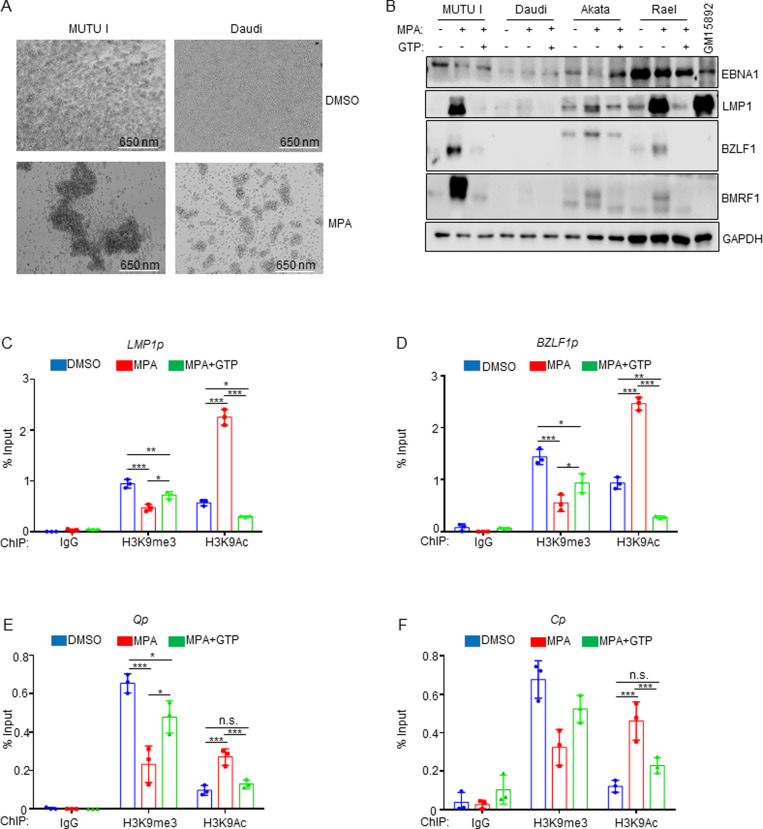
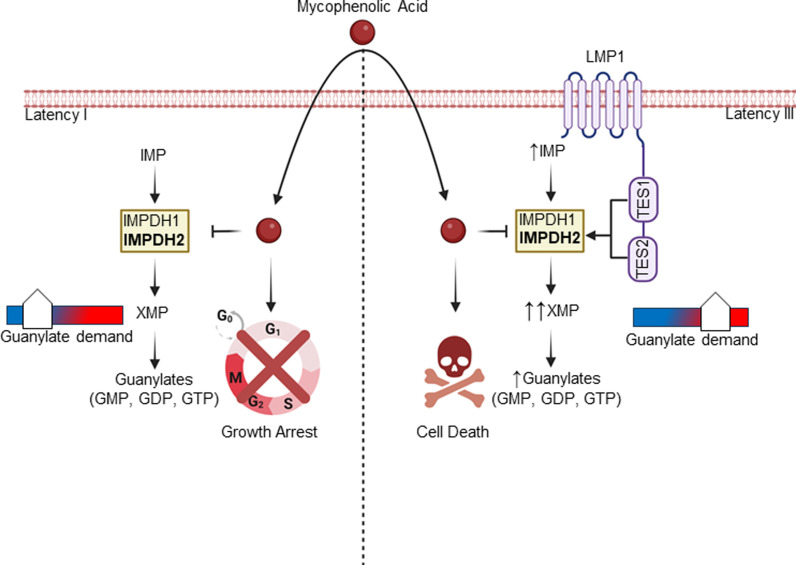
Epstein-Barr virus (EBV) is associated with multiple types of cancers, many of which express the viral oncoprotein Latent Membrane Protein 1 (LMP1). LMP1 contributes to both epithelial and B-cell transformation. Although metabolism reprogramming is a cancer hallmark, much remains to be learned about how LMP1 alters lymphocyte oncometabolism. To gain insights into key B-cell metabolic pathways subverted by LMP1, we performed systematic metabolomic analyses on B cells with conditional LMP1 expression. This approach highlighted that LMP highly induces de novo purine biosynthesis, with xanthosine-5-P (XMP) as one of the most highly LMP1-upregulated metabolites. Consequently, IMPDH inhibition by mycophenolic acid (MPA) triggered death of LMP1-expressing EBV-transformed lymphoblastoid cell lines (LCL), a key model for EBV-driven immunoblastic lymphomas. Whereas MPA instead caused growth arrest of Burkitt lymphoma cells with the EBV latency I program, conditional LMP1 expression triggered their death, and this phenotype was rescuable by guanosine triphosphate (GTP) supplementation, implicating LMP1 as a key driver of B-cell GTP biosynthesis. Although both IMPDH isozymes are expressed in LCLs, only IMPDH2 was critical for LCL survival, whereas both contributed to proliferation of Burkitt cells with the EBV latency I program. Both LMP1 C-terminal cytoplasmic tail domains critical for primary human B-cell transformation were important for XMP production, and each contributed to LMP1-driven Burkitt cell sensitivity to MPA. Metabolomic analyses further highlighted roles of NF-kB, mitogen activated kinase, and protein kinase C downstream of LMP1 in support of XMP abundance. Of these, only protein kinase C activity was important for supporting GTP levels in LMP1 expressing Burkitt cells. MPA also de-repressed EBV lytic antigens, including LMP1 itself in latency I Burkitt cells, highlighting crosstalk between the purine biosynthesis pathway and the EBV epigenome. These results suggest novel oncometabolism-based therapeutic approaches to LMP1-driven lymphomas.
Copyright: © 2025 Burton et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
Epstein-Barr Virus Latent Membrane Protein 1 Subverts IMPDH pathways to drive B-cell oncometabolism.bioRxiv [Preprint]. 2024 Nov 8:2024.11.07.622457. doi: 10.1101/2024.11.07.622457. bioRxiv. 2024. Update in: PLoS Pathog. 2025 May 14;21(5):e1013092. doi: 10.1371/journal.ppat.1013092. PMID: 39574729 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
