

Virus and cell specific HMGB1 secretion and subepithelial infiltrate formation in adenovirus keratitis
- PMID: 40367285
- PMCID: PMC12101768
- DOI: 10.1371/journal.ppat.1013184
Virus and cell specific HMGB1 secretion and subepithelial infiltrate formation in adenovirus keratitis
Abstract
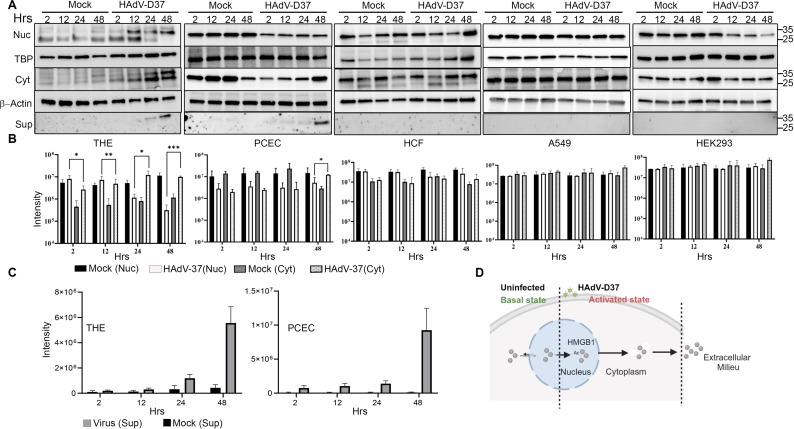
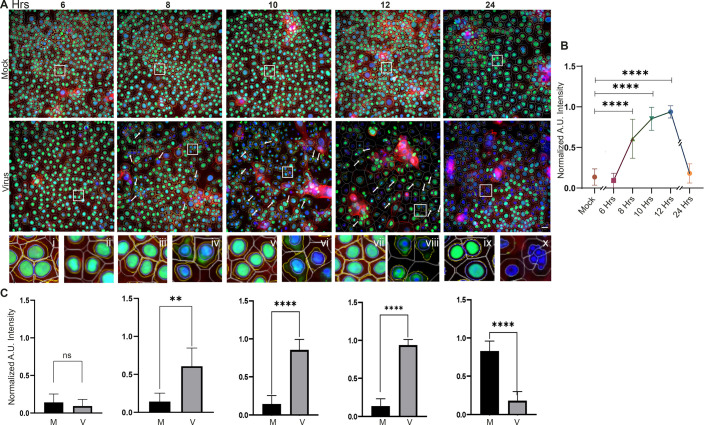
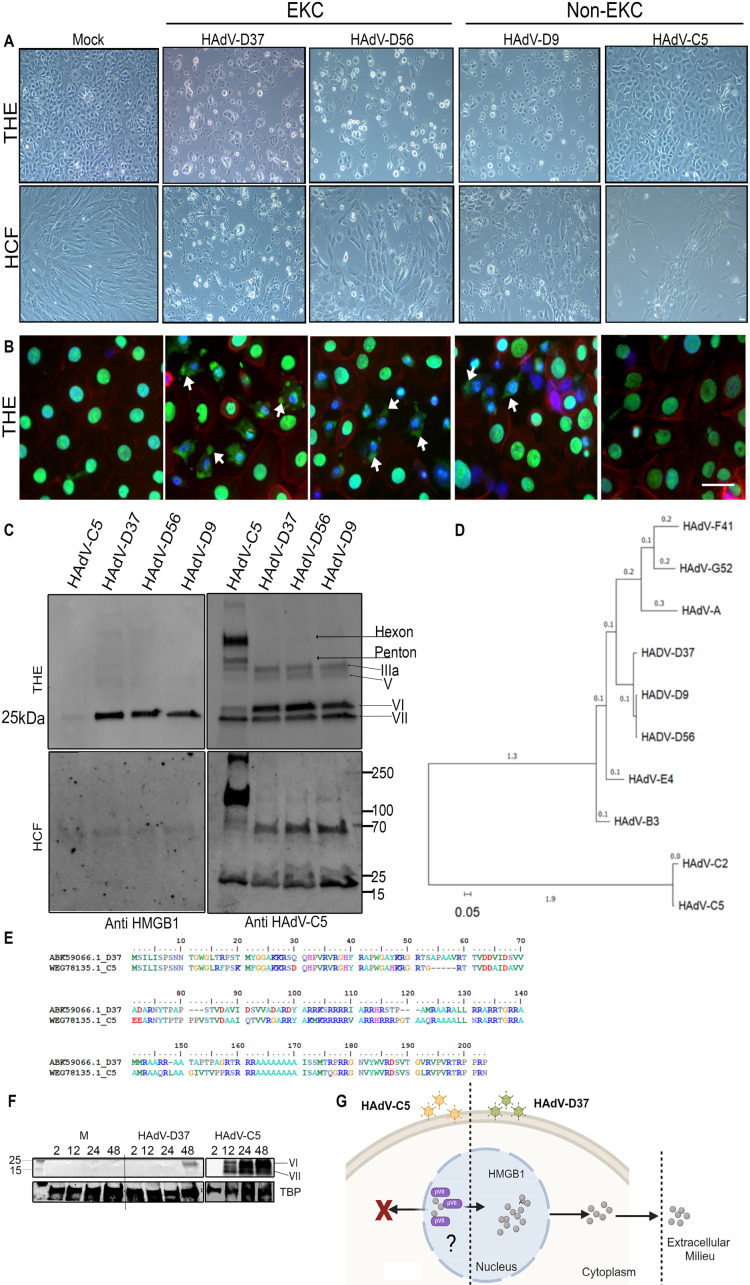
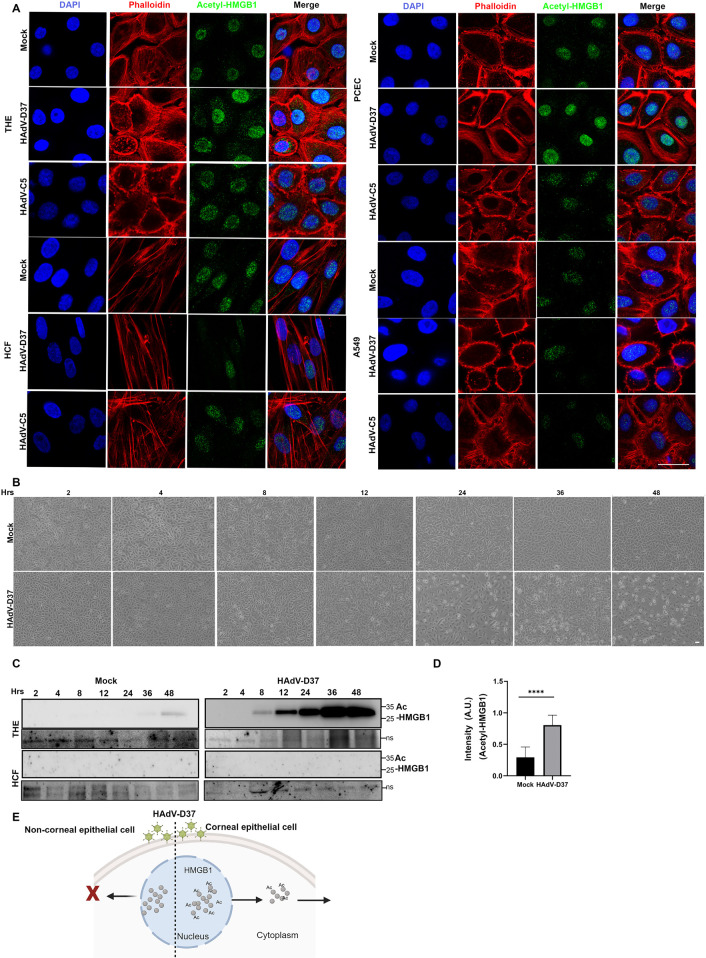
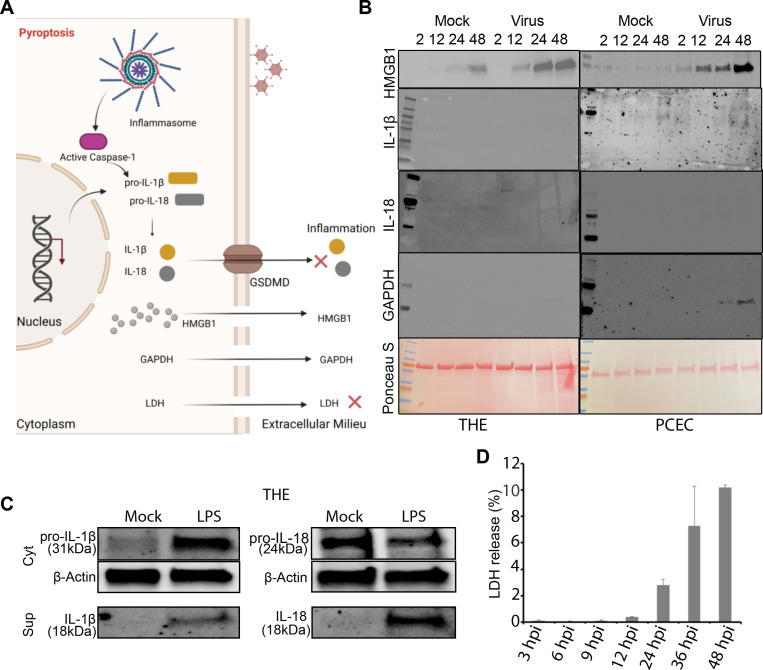
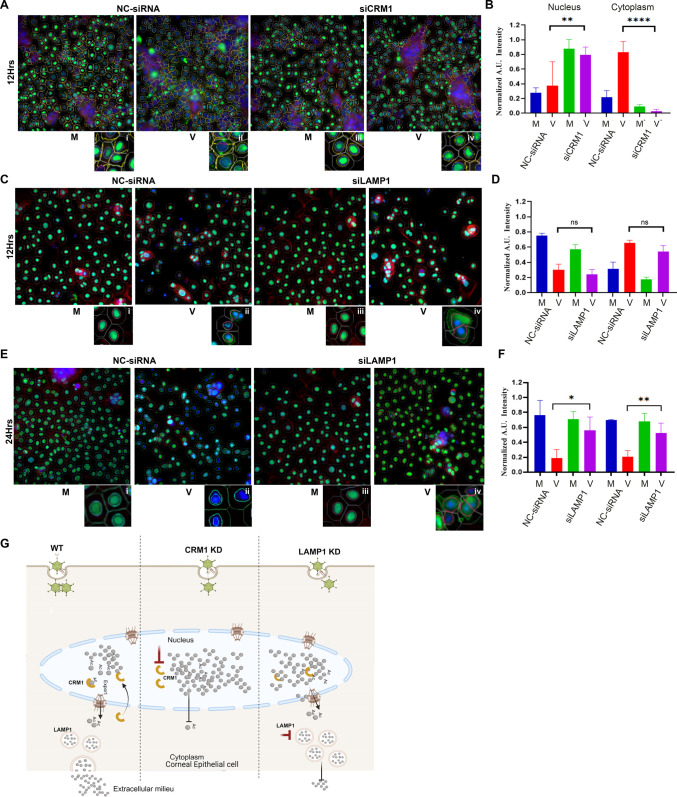
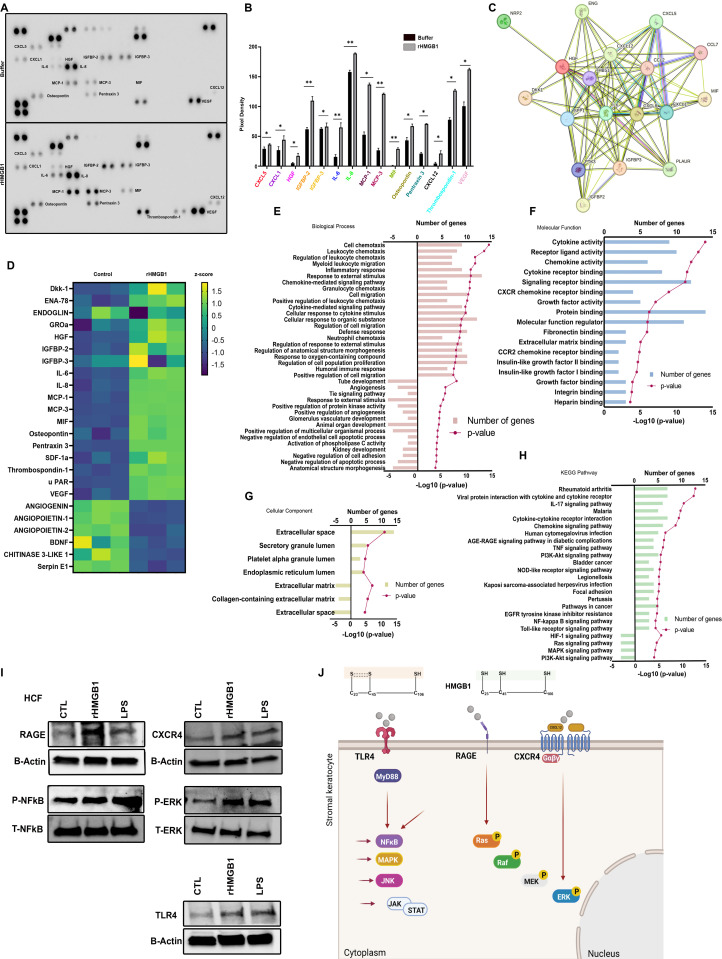
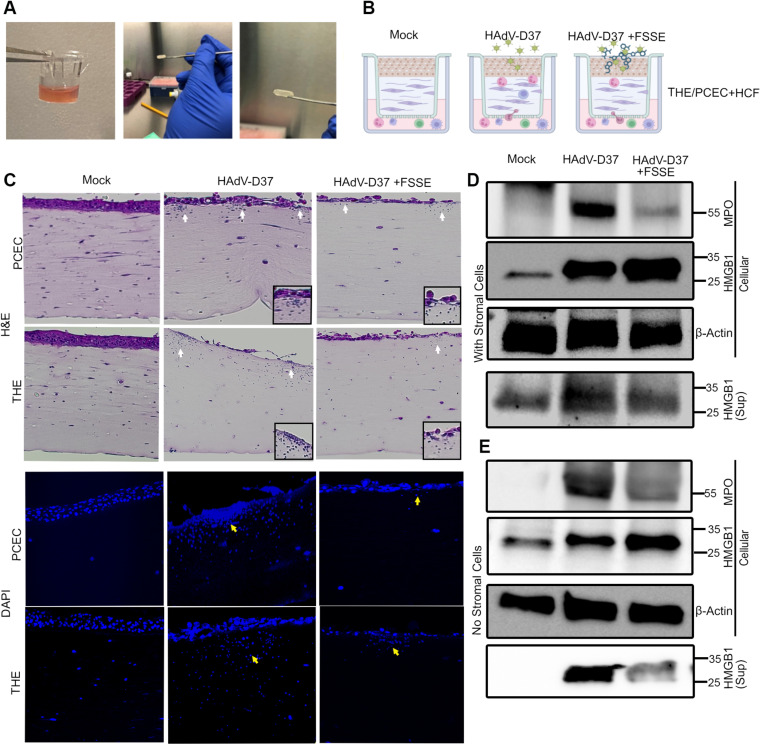
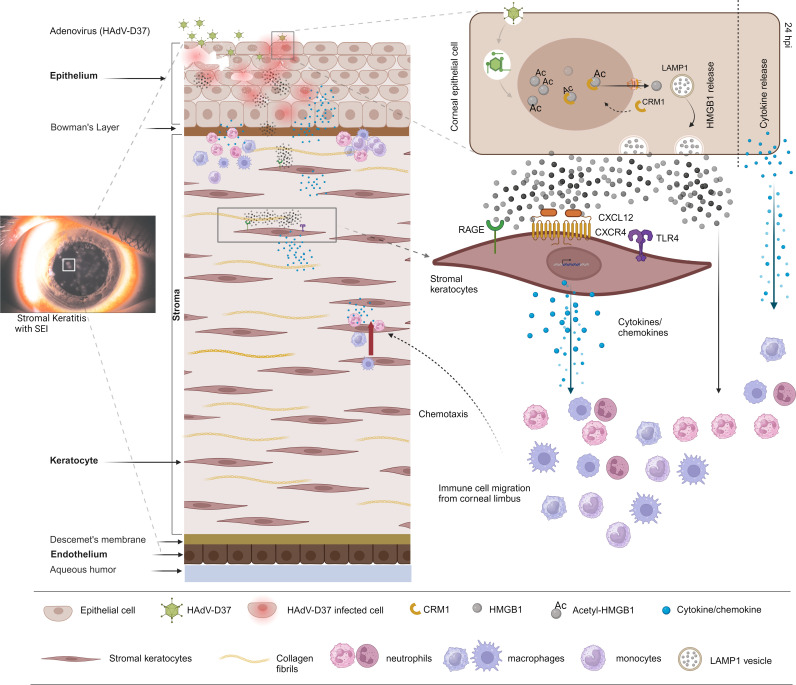
A highly contagious infection caused by human adenovirus species D (HAdV-D), epidemic keratoconjunctivitis (EKC) results in corneal subepithelial infiltration (SEI) by leukocytes, the hallmark of the infection. To date, the pathogenesis of corneal SEI formation in EKC is unresolved. HMGB1 (high-mobility group box 1 protein) is an alarmin expressed in response to infection and a marker of sepsis. Earlier studies using a different adenovirus species, HAdV-C, showed retention of HMGB1 in the infected cell nucleus by adenovirus protein VII, enabling immune evasion. Here, using HAdV-D we show cell-specific HMGB1 secretion by infected cells, and provide an HAdV-D specific mechanism for SEI formation in EKC. HMGB1 was secreted only upon infection of human corneal epithelial cells, not from other cell types, and only upon infection by HAdV-D types associated with EKC. Acetylated HMGB1 translocation from the nucleus to the cytoplasm, then to the extracellular milieu, was tightly controlled by CRM1 and LAMP1, respectively. Primary stromal cells when stimulated by rHMGB1 expressed proinflammatory chemokines. In a novel 3D culture system in tune with the architecture of the cornea, HMGB1 released by infected corneal epithelial cells induced leukocytic infiltrates either directly and/or indirectly via stimulated stromal cells, which together explains SEI formation in EKC.
Copyright: © 2025 Saha et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
Virus and Cell Specific HMGB1 Secretion and Subepithelial Infiltrate Formation in Adenovirus Keratitis.bioRxiv [Preprint]. 2025 Jan 7:2025.01.07.631509. doi: 10.1101/2025.01.07.631509. bioRxiv. 2025. Update in: PLoS Pathog. 2025 May 14;21(5):e1013184. doi: 10.1371/journal.ppat.1013184. PMID: 39829903 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
