

Spatial transcriptomics reveals human cortical layer and area specification
- PMID: 40369074
- PMCID: PMC12328223
- DOI: 10.1038/s41586-025-09010-1
Spatial transcriptomics reveals human cortical layer and area specification
Abstract
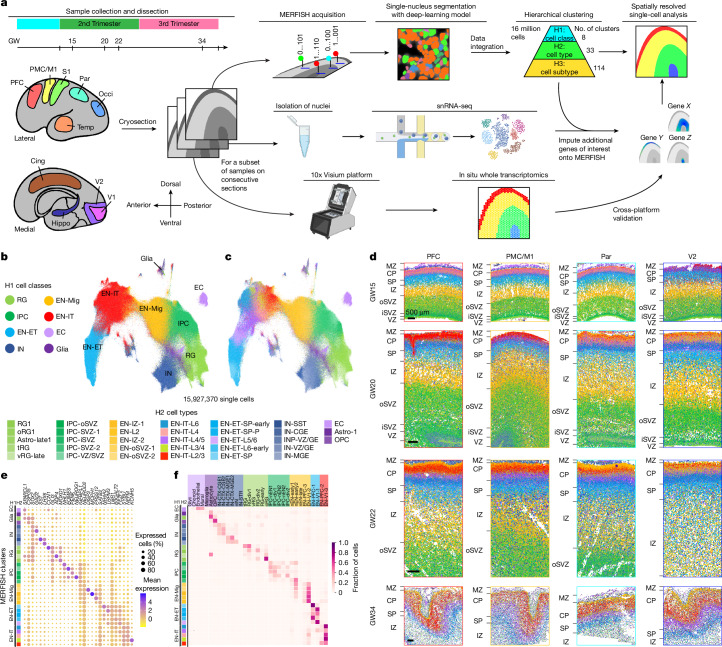
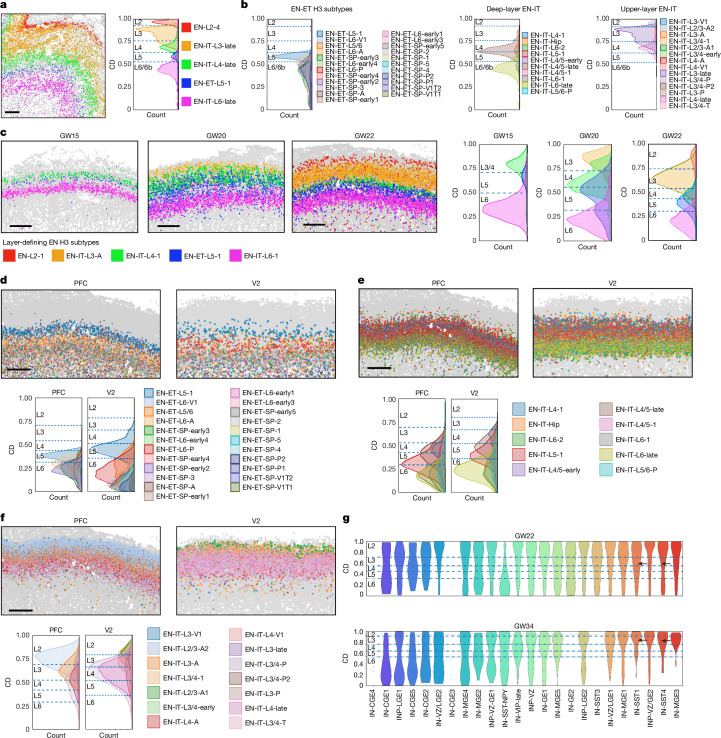
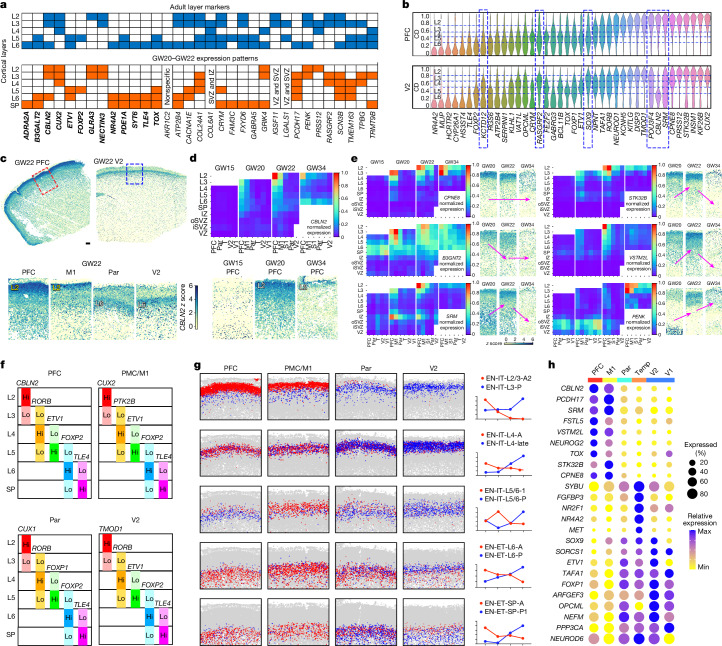
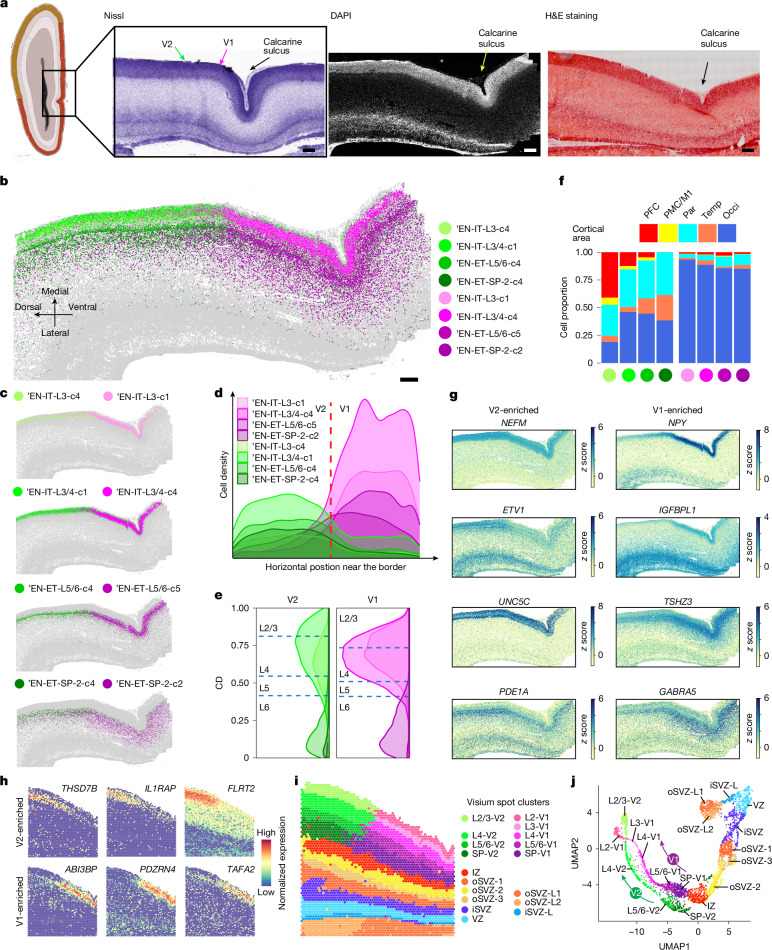
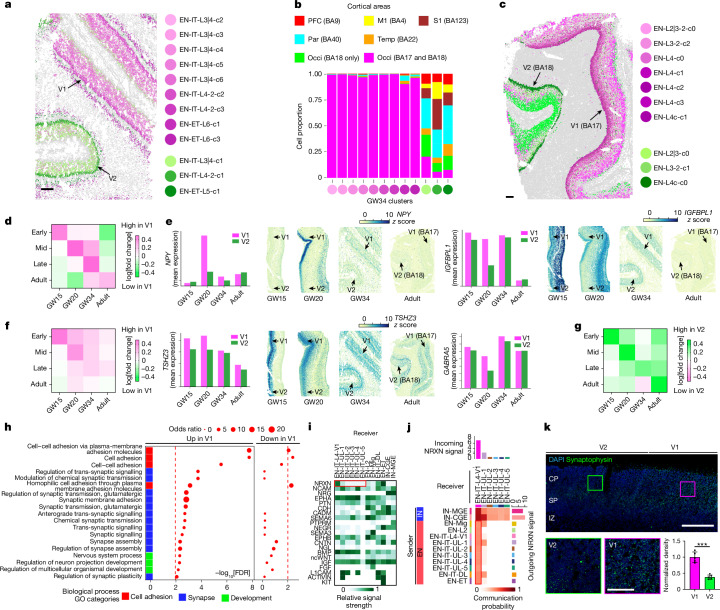
The human cerebral cortex is composed of six layers and dozens of areas that are molecularly and structurally distinct1-4. Although single-cell transcriptomic studies have advanced the molecular characterization of human cortical development, a substantial gap exists owing to the loss of spatial context during cell dissociation5-8. Here we used multiplexed error-robust fluorescence in situ hybridization (MERFISH)9, augmented with deep-learning-based nucleus segmentation, to examine the molecular, cellular and cytoarchitectural development of the human fetal cortex with spatially resolved single-cell resolution. Our extensive spatial atlas, encompassing more than 18 million single cells, spans eight cortical areas across seven developmental time points. We uncovered the early establishment of the six-layer structure, identifiable by the laminar distribution of excitatory neuron subtypes, 3 months before the emergence of cytoarchitectural layers. Notably, we discovered two distinct modes of cortical areal specification during mid-gestation: (1) a continuous, gradual transition observed across most cortical areas along the anterior-posterior axis and (2) a discrete, abrupt boundary specifically identified between the primary (V1) and secondary (V2) visual cortices as early as gestational week 20. This sharp binary transition in V1-V2 neuronal subtypes challenges the notion that mid-gestation cortical arealization involves only gradient-like transitions6,10. Furthermore, integrating single-nucleus RNA sequencing with MERFISH revealed an early upregulation of synaptogenesis in V1-specific layer 4 neurons. Collectively, our findings underscore the crucial role of spatial relationships in determining the molecular specification of cortical layers and areas. This study establishes a spatially resolved single-cell analysis paradigm and paves the way for the construction of a comprehensive developmental atlas of the human brain.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: C.A.W. has stock ownership in Maze Therapeutics and Mosaica Therapeutics, and is a paid consultant for Third Rock Ventures and Flagship Pioneering. M.L. receives research funding from Biogen, unrelated to the current article. M.L. is a co-founder of OmicPath AI. The remaining authors declare no competing interests.
Figures


















References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
