

Taurine from tumour niche drives glycolysis to promote leukaemogenesis
- PMID: 40369079
- PMCID: PMC12328231
- DOI: 10.1038/s41586-025-09018-7
Taurine from tumour niche drives glycolysis to promote leukaemogenesis
Abstract
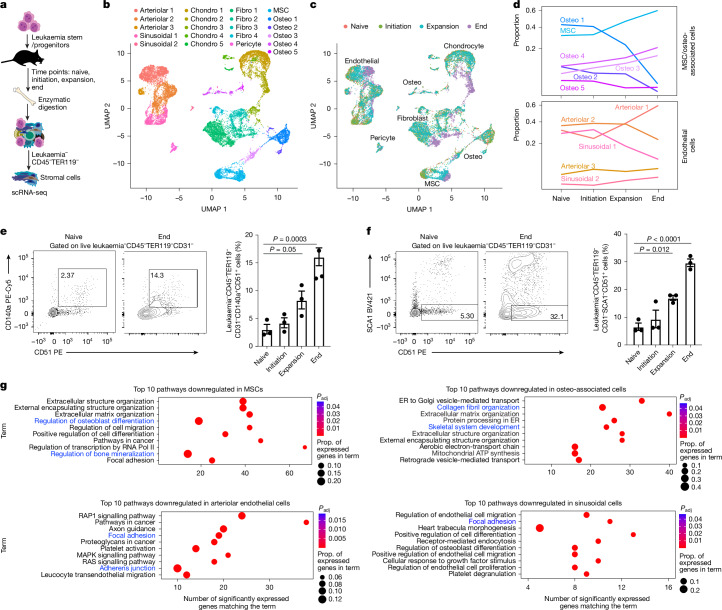
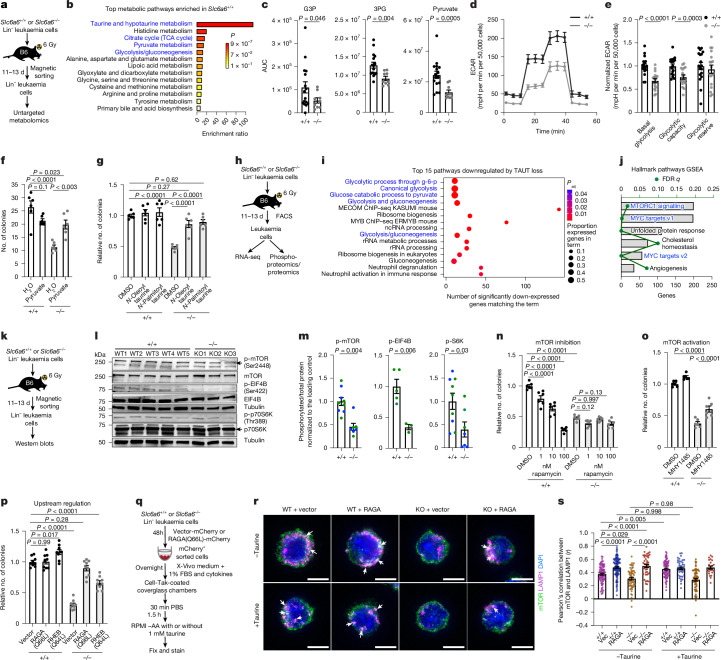
Signals from the microenvironment are known to be critical for development, stem cell self-renewal and oncogenic progression. Although some niche-driven signals that promote cancer progression have been identified1-5, concerted efforts to map disease-relevant microenvironmental ligands of cancer stem cell receptors have been lacking. Here, we use temporal single-cell RNA-sequencing (scRNA-seq) to identify molecular cues from the bone marrow stromal niche that engage leukaemia stem-enriched cells (LSCs) during oncogenic progression. We integrate these data with our human LSC RNA-seq and in vivo CRISPR screen of LSC dependencies6 to identify LSC-niche interactions that are essential for leukaemogenesis. These analyses identify the taurine-taurine transporter (TAUT) axis as a critical dependency of aggressive myeloid leukaemias. We find that cysteine dioxygenase type 1 (CDO1)-driven taurine biosynthesis is restricted to osteolineage cells, and increases during myeloid disease progression. Blocking CDO1 expression in osteolineage cells impairs LSC growth and improves survival outcomes. Using TAUT genetic loss-of-function mouse models and patient-derived acute myeloid leukaemia (AML) cells, we show that TAUT inhibition significantly impairs in vivo myeloid leukaemia progression. Consistent with elevated TAUT expression in venetoclax-resistant AML, TAUT inhibition synergizes with venetoclax to block the growth of primary human AML cells. Mechanistically, our multiomic approaches indicate that the loss of taurine uptake inhibits RAG-GTP dependent mTOR activation and downstream glycolysis. Collectively, our work establishes the temporal landscape of stromal signals during leukaemia progression and identifies taurine as a key regulator of myeloid malignancies.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures















References
-
- Orimo, A. et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell121, 335–348 (2005). - PubMed
-
- Vermeulen, L. et al. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat. Cell Biol.12, 468–476 (2010). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
