

CDK5-triggered G6PD phosphorylation at threonine 91 facilitating redox homeostasis reveals a vulnerability in breast cancer
- PMID: 40370560
- PMCID: PMC12069116
- DOI: 10.1016/j.apsb.2024.12.019
CDK5-triggered G6PD phosphorylation at threonine 91 facilitating redox homeostasis reveals a vulnerability in breast cancer
Abstract
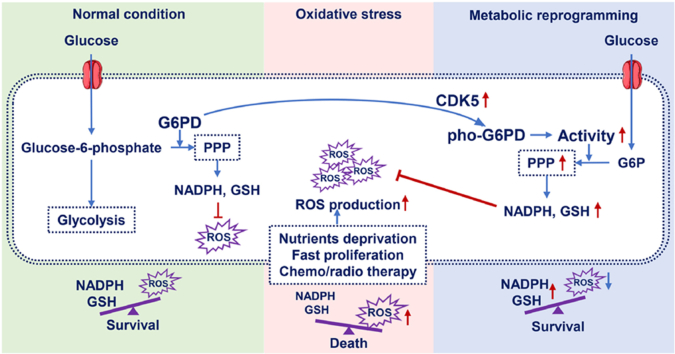
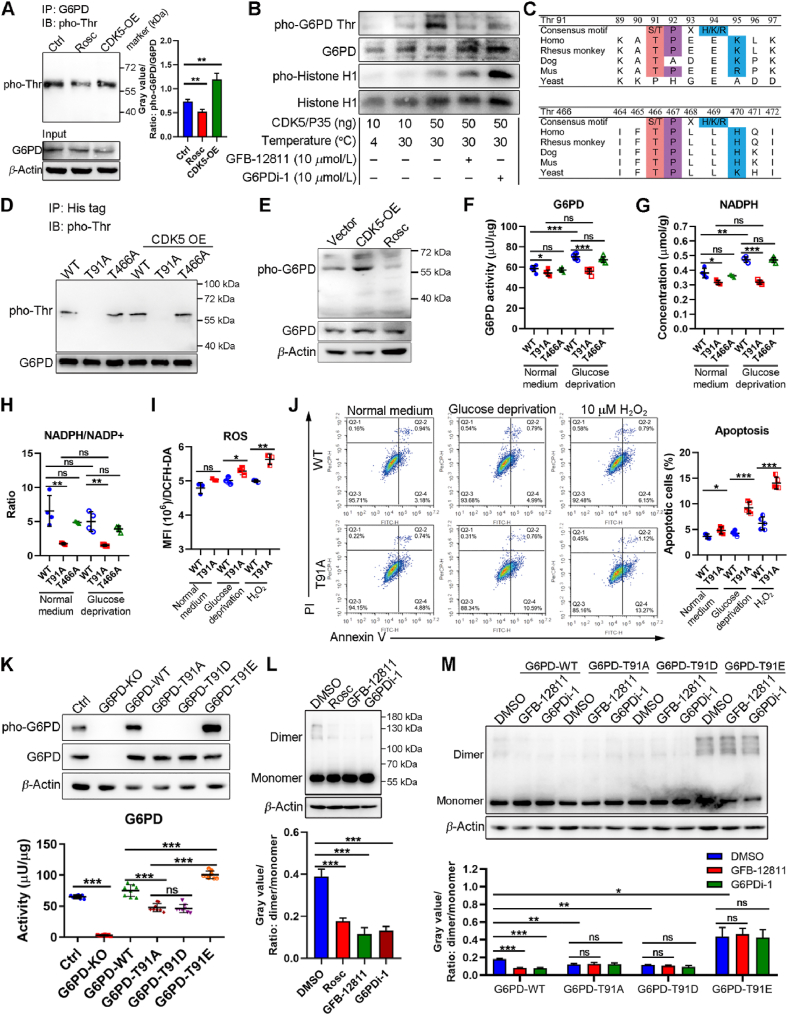
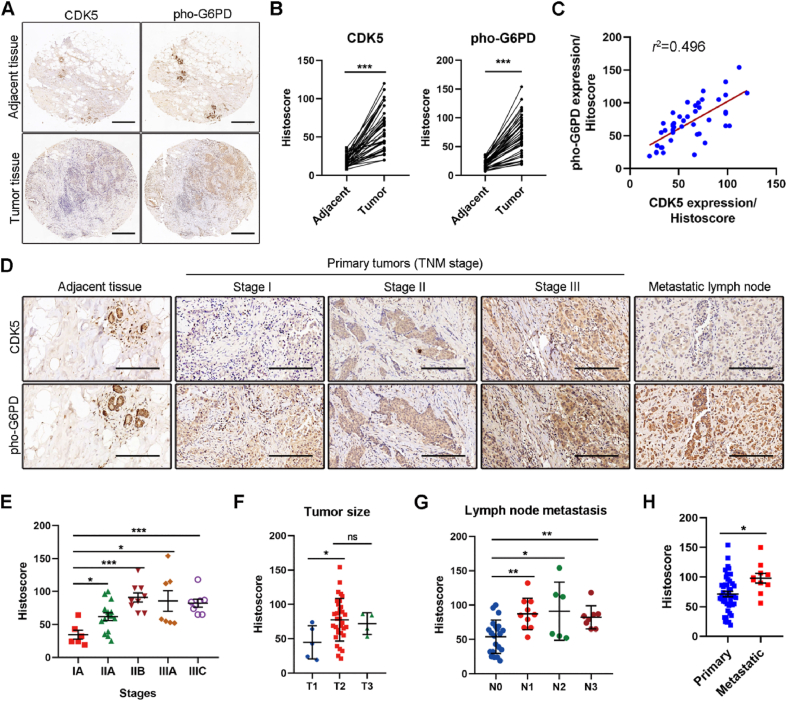
Glucose-6-phosphate dehydrogenase (G6PD), the first rate-limiting enzyme of the pentose phosphate pathway (PPP), is aberrantly activated in multiple types of human cancers, governing the progression of tumor cells as well as the efficacy of anticancer therapy. Here, we discovered that cyclin-dependent kinase 5 (CDK5) rewired glucose metabolism from glycolysis to PPP in breast cancer (BC) cells by activating G6PD to keep intracellular redox homeostasis under oxidative stress. Mechanistically, CDK5-phosphorylated G6PD at Thr-91 facilitated the assembly of inactive monomers of G6PD into active dimers. More importantly, CDK5-induced pho-G6PD was explicitly observed specifically in tumor tissues in human BC specimens. Pharmacological inhibition of CDK5 remarkably abrogated G6PD phosphorylation, attenuated tumor growth and metastasis, and synergistically sensitized BC cells to poly-ADP-ribose polymerase (PARP) inhibitor Olaparib, in xenograft mouse models. Collectively, our results establish the crucial role of CDK5-mediated phosphorylation of G6PD in BC growth and metastasis and provide a therapeutic regimen for BC treatment.
Keywords: Breast cancer; CDK5; Drug resistance; Glucose-6-phosphoate dehydrogenase; Intracellular redox homeostasis; Isotopomer spectral analysis; Olaparib; Pentose phosphate pathway.
© 2025 The Authors.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





Similar articles
-
Rapid phosphorylation of glucose-6-phosphate dehydrogenase by casein kinase 2 sustains redox homeostasis under ionizing radiation.Redox Biol. 2023 Sep;65:102810. doi: 10.1016/j.redox.2023.102810. Epub 2023 Jul 7. Redox Biol. 2023. PMID: 37478541 Free PMC article.
-
HuaChanSu suppresses the growth of hepatocellular carcinoma cells by interfering with pentose phosphate pathway through down-regulation of G6PD enzyme activity and expression.Heliyon. 2024 Jan 26;10(3):e25144. doi: 10.1016/j.heliyon.2024.e25144. eCollection 2024 Feb 15. Heliyon. 2024. PMID: 38322888 Free PMC article.
-
The Emerging Roles of the Metabolic Regulator G6PD in Human Cancers.Int J Mol Sci. 2023 Dec 7;24(24):17238. doi: 10.3390/ijms242417238. Int J Mol Sci. 2023. PMID: 38139067 Free PMC article. Review.
-
Glucose-6-phosphate dehydrogenase regulates mitophagy by maintaining PINK1 stability.Life Metab. 2024 Dec 13;4(1):loae040. doi: 10.1093/lifemeta/loae040. eCollection 2025 Feb. Life Metab. 2024. PMID: 39872984 Free PMC article.
-
Recent findings in the regulation of G6PD and its role in diseases.Front Pharmacol. 2022 Aug 24;13:932154. doi: 10.3389/fphar.2022.932154. eCollection 2022. Front Pharmacol. 2022. PMID: 36091812 Free PMC article. Review.
References
-
- Hanahan D., Weinberg Robert A. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
-
- de Mas I.M., Aguilar E., Jayaraman A., Polat I.H., Martin-Bernabe A., Bharat R., et al. Cancer cell metabolism as new targets for novel designed therapies. Future Med Chem. 2014;6:1791–1810. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
