

Spatiotemporal Dynamics of Non-Ecological Speciation in Rubyspot Damselflies (Hetaerina spp.)
- PMID: 40371671
- PMCID: PMC12143376
- DOI: 10.1111/mec.17797
Spatiotemporal Dynamics of Non-Ecological Speciation in Rubyspot Damselflies (Hetaerina spp.)
Abstract
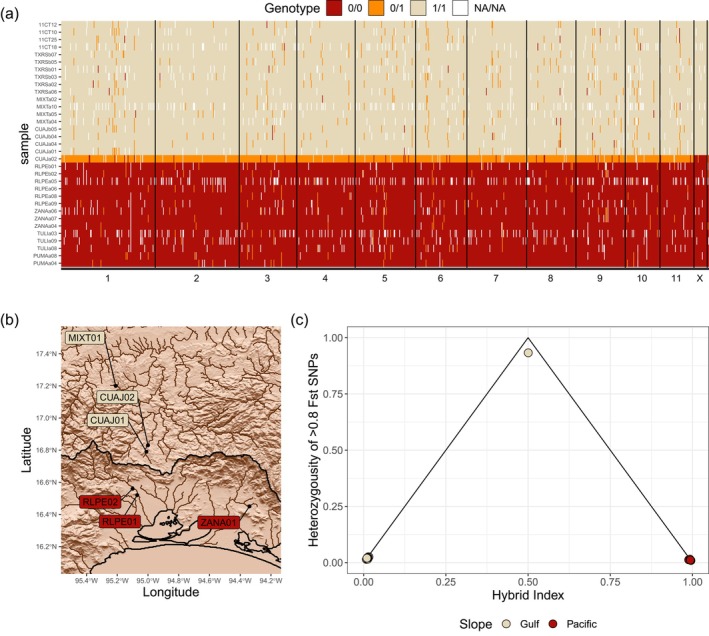
Non-ecological speciation is a common mode of speciation, which occurs when allopatric lineages diverge in the absence of pronounced ecological differences. Yet, relative to other speciation mechanisms, non-ecological speciation remains understudied. Numerous damselfly clades are characterised as non-adaptive radiations (the result of several rounds of non-ecological speciation without subsequent divergence), but there are few damselfly lineages for which we have a detailed understanding of the spatiotemporal dynamics of divergence. Recent phylogeographic analyses demonstrate that American rubyspot damselflies (Hetaerina americana sensu lato) actually comprise at least two cryptic lineages that coexist sympatrically across most of Mexico. To broaden our understanding of the dynamics of diversification to other rubyspot lineages, we investigated the phylogeographic history of smoky rubyspot damselflies (Hetaerina titia) using genomic data collected across Central and North America. Unexpectedly, we found evidence of reproductive isolation between the highly genetically differentiated Pacific and Atlantic lineages of H. titia in a narrow secondary contact zone on the Isthmus of Tehuantepec, Mexico. We then fit models of historical demography to both H. americana sensu lato and H. titia to place these comparisons in a temporal context. Our findings indicate that Pacific and Atlantic lineages of H. titia split more recently than the broadly sympatric lineages within H. americana sensu lato, supporting key assumptions of the non-ecological speciation model and demonstrating that these two pairs of sister lineages are at different stages of the speciation cycle.
La especiación no‐ecológica es un modo común de especiación que ocurre cuando los linajes alopátricos divergen sin diferencias ecológicas pronunciadas. Sin embargo, comparado con otros mecanismos de especiación, la especiación no‐ecológica sigue siendo poco estudiada. Varios clados de caballitos del diablo se han caracterizado como radiaciones no‐adaptativas (es decir, el resultado de varios ciclos de especiación no‐ecológica sin divergencia posterior), pero hay pocos ejemplos de caballitos del diablo para los cuales tenemos un conocimiento detallado de la dinámica espaciotemporal de la divergencia. Análisis filogeográficos recientes demuestran que el caballito escarlata común (Hetaerina americana sensu lato) en realidad consiste en al menos dos linajes crípticos que coexisten simpátricamente a lo largo de México. Para ampliar nuestra comprensión de la dinámica de la diversificación a otros linajes de los caballitos escarlatas, investigamos patrones filogeográficos del caballito escarlata de alas ahumadas (Hetaerina titia) utilizando datos genómicos de individuos de América Central y del Norte. Inesperadamente, encontramos evidencia de que las poblaciones altamente diferenciadas genéticamente del Pacífico y el Atlántico de H. titia exhiben aislamiento reproductivo en un punto de contacto secundario en el Istmo de Tehuantepec en México. Además, modelamos la demografía histórica tanto de H. americana sensu lato como de H. titia para compararlas en un contexto temporal. Descubrimos que los linajes del Pacífico y el Atlántico de H. titia se dividieron más recientemente que los linajes hermanos y ampliamente simpátricos de H. americana sensu lato, lo cual apoya las suposiciones claves del modelo de especiación no‐ecológica y demuestra que estos dos pares de linajes hermanos se encuentran en diferentes etapas del ciclo de especiación no‐ecológica.
Keywords: Odonata; Zygoptera; ddRAD; population genomics; speciation.
© 2025 The Author(s). Molecular Ecology published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





Similar articles
-
A chromosome-level genome assembly for the smoky rubyspot damselfly (Hetaerina titia).J Hered. 2024 Feb 3;115(1):103-111. doi: 10.1093/jhered/esad070. J Hered. 2024. PMID: 37988159 Free PMC article.
-
Complex evolutionary history of the American Rubyspot damselfly, Hetaerina americana (Odonata): Evidence of cryptic speciation.Mol Phylogenet Evol. 2019 Oct;139:106536. doi: 10.1016/j.ympev.2019.106536. Epub 2019 Jun 15. Mol Phylogenet Evol. 2019. PMID: 31212083
-
Phylogeography of a cryptic speciation continuum in Eurasian spadefoot toads (Pelobates).Mol Ecol. 2019 Jul;28(13):3257-3270. doi: 10.1111/mec.15133. Epub 2019 Jun 28. Mol Ecol. 2019. PMID: 31254307
-
Reference genome for the American rubyspot damselfly, Hetaerina americana.J Hered. 2023 Jun 22;114(4):385-394. doi: 10.1093/jhered/esad031. J Hered. 2023. PMID: 37195415 Free PMC article.
-
Seasonal polyphenism in wing coloration affects species recognition in rubyspot damselflies (Hetaerina spp.).J Evol Biol. 2015 Aug;28(8):1439-52. doi: 10.1111/jeb.12665. Epub 2015 Jun 11. J Evol Biol. 2015. PMID: 26033550
References
-
- Anderson, C. N. , and Grether G. F.. 2011. “Multiple Routes to Reduced Interspecific Territorial Fighting in Hetaerina Damselflies.” Behavioral Ecology 22, no. 3: 527–534. 10.1093/beheco/arr013. - DOI
-
- Anderson, S. A. , López‐Fernández H., and Weir J. T.. 2023. “Ecology and the Origin of Nonephemeral Species.” American Naturalist 201, no. 5: 619–638. - PubMed
-
- Barrier, E. , Velasquillo L., Chavez M., and Gaulon R.. 1998. “Neotectonic Evolution of the Isthmus of Tehuantepec (Southeastern Mexico).” Tectonophysics 287, no. 1: 77–96. 10.1016/S0040-1951(98)80062-0. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
