

Intrinsically Disordered Regions Define Unique Protein Interaction Networks in CHD Family Remodelers
- PMID: 40372282
- PMCID: PMC12080455
- DOI: 10.1096/fj.202402808RR
Intrinsically Disordered Regions Define Unique Protein Interaction Networks in CHD Family Remodelers
Abstract
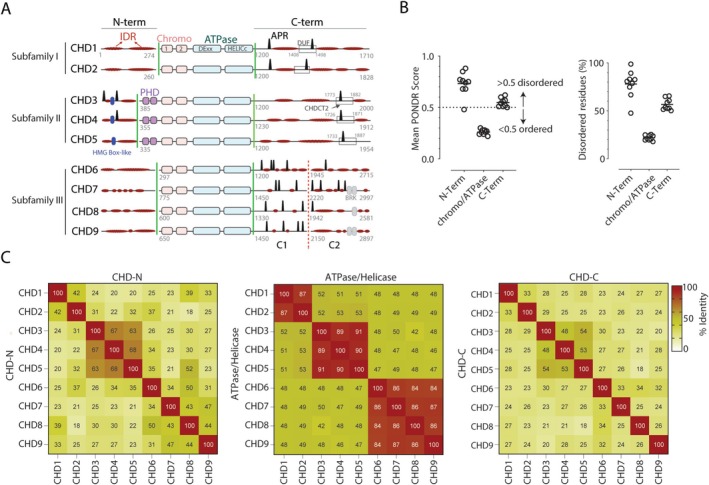
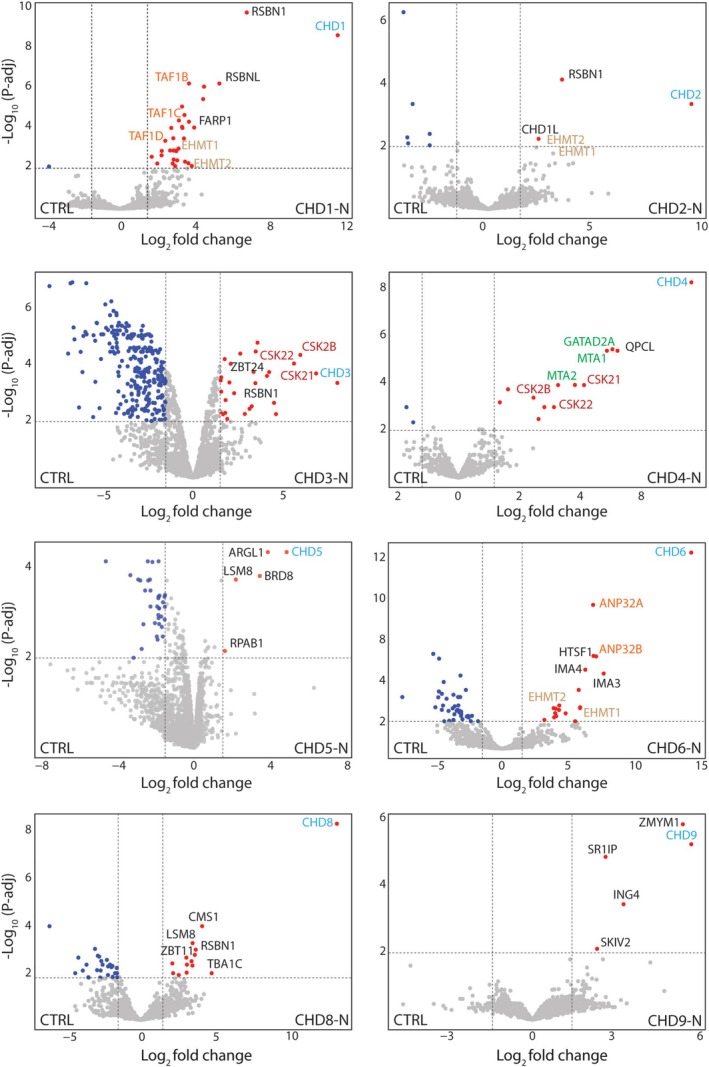
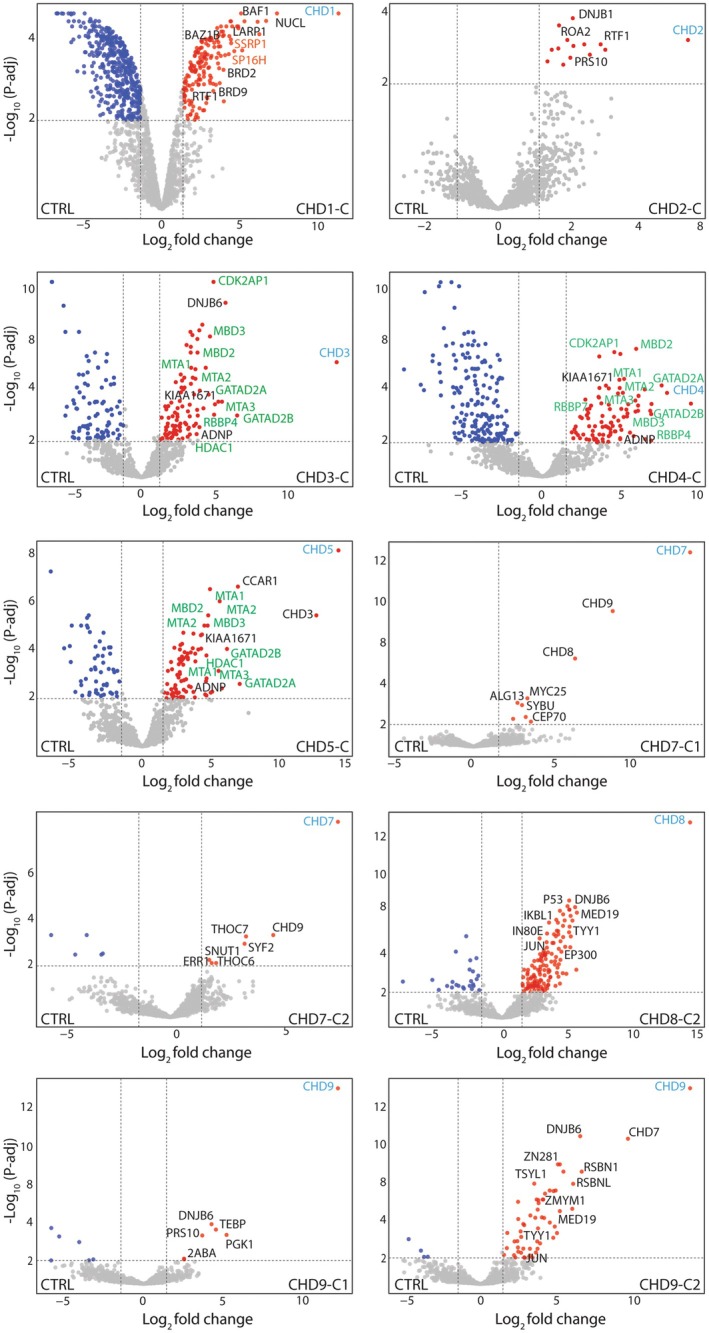
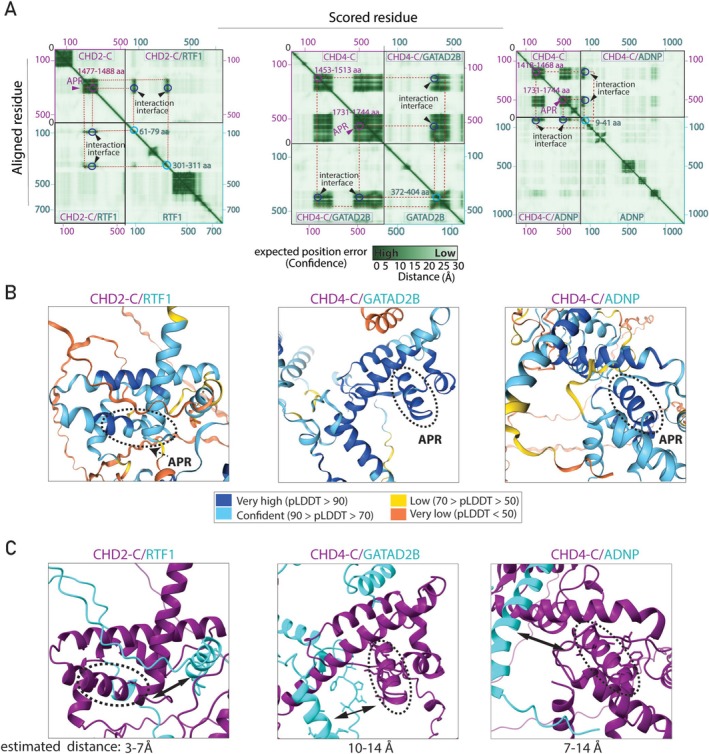
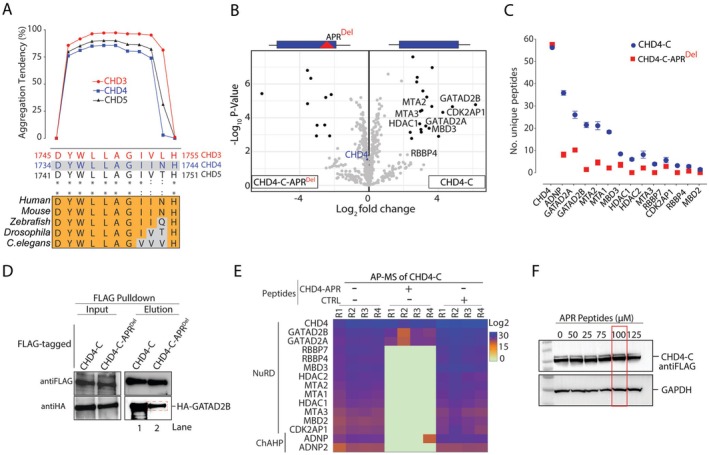
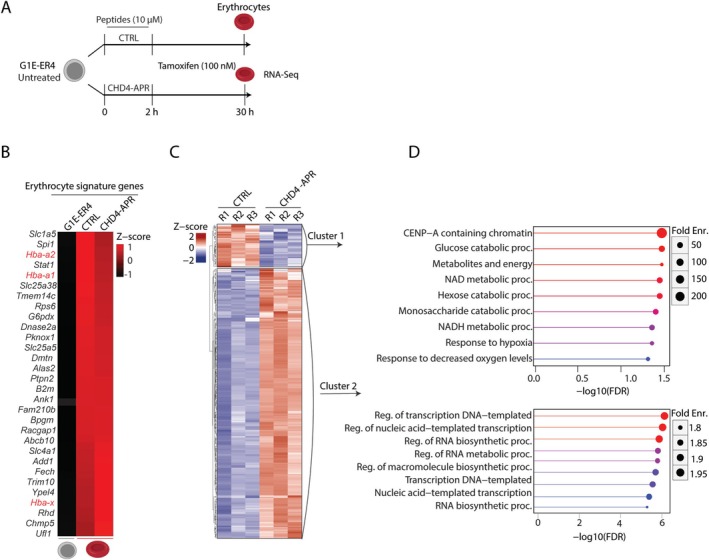
Chromodomain helicase DNA-binding (CHD) enzymes play a pivotal role in genome regulation. They possess highly conserved ATPase domains flanked by poorly characterized and intrinsically disordered N- and C-termini. Using mass spectrometry, we identify dozens of novel protein-protein interactions (PPIs) within the N- and C-termini of human CHD family members. We also define a highly conserved aggregation-prone region (APR) within the C-terminus of CHD4 which is critical for its interaction with the nucleosome remodeling and deacetylase (NuRD), as well as ChAHP (CHD4, activity-dependent neuroprotective protein (ADNP), and HP1γ) complexes. Further analysis reveals a regulatory role for the CHD4 APR in gene transcription during erythrocyte formation. Our results highlight that the N- and C-termini of CHD chromatin remodelers shape protein interaction networks that drive unique transcriptional programs.
Keywords: DNA helicase; aggregation‐prone regions; chromatin remodelers; intrinsically disordered regions; protein–protein interactions; transcription.
© 2025 The Author(s). The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Unique protein interaction networks define the chromatin remodelling module of the NuRD complex.FEBS J. 2022 Jan;289(1):199-214. doi: 10.1111/febs.16112. Epub 2021 Jul 31. FEBS J. 2022. PMID: 34231305 Free PMC article.
-
CHD4 Is a Peripheral Component of the Nucleosome Remodeling and Deacetylase Complex.J Biol Chem. 2016 Jul 22;291(30):15853-66. doi: 10.1074/jbc.M115.707018. Epub 2016 May 27. J Biol Chem. 2016. PMID: 27235397 Free PMC article.
-
An intrinsically disordered region of methyl-CpG binding domain protein 2 (MBD2) recruits the histone deacetylase core of the NuRD complex.Nucleic Acids Res. 2015 Mar 31;43(6):3100-13. doi: 10.1093/nar/gkv168. Epub 2015 Mar 9. Nucleic Acids Res. 2015. PMID: 25753662 Free PMC article.
-
Chromatin Remodulator CHD4: A Potential Target for Cancer Interception.Genes (Basel). 2025 Feb 15;16(2):225. doi: 10.3390/genes16020225. Genes (Basel). 2025. PMID: 40004553 Free PMC article. Review.
-
Chromodomain helicase DNA-binding 4 (CHD4) regulates early B cell identity and V(D)J recombination.Immunol Rev. 2022 Jan;305(1):29-42. doi: 10.1111/imr.13054. Epub 2021 Dec 20. Immunol Rev. 2022. PMID: 34927255 Free PMC article. Review.
Cited by
-
A direct interaction between the Chd1 CHCT domain and Rtf1 controls Chd1 distribution and nucleosome positioning on active genes.bioRxiv [Preprint]. 2024 Dec 6:2024.12.06.627179. doi: 10.1101/2024.12.06.627179. bioRxiv. 2024. PMID: 39677735 Free PMC article. Preprint.
References
-
- Bednar J., Horowitz R. A., Grigoryev S. A., et al., “Nucleosomes, Linker DNA, and Linker Histone Form a Unique Structural Motif That Directs the Higher‐Order Folding and Compaction of Chromatin,” Proceedings of the National Academy of Sciences of the United States of America 95 (1998): 14173–14178. - PMC - PubMed
-
- Saha A., Wittmeyer J., and Cairns B. R., “Chromatin Remodelling: The Industrial Revolution of DNA Around Histones,” Nature Reviews. Molecular Cell Biology 7 (2006): 437–447. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
