

Gfi1 controls the formation of effector-like CD8+ T cells during chronic infection and cancer
- PMID: 40374625
- PMCID: PMC12081725
- DOI: 10.1038/s41467-025-59784-1
Gfi1 controls the formation of effector-like CD8+ T cells during chronic infection and cancer
Abstract
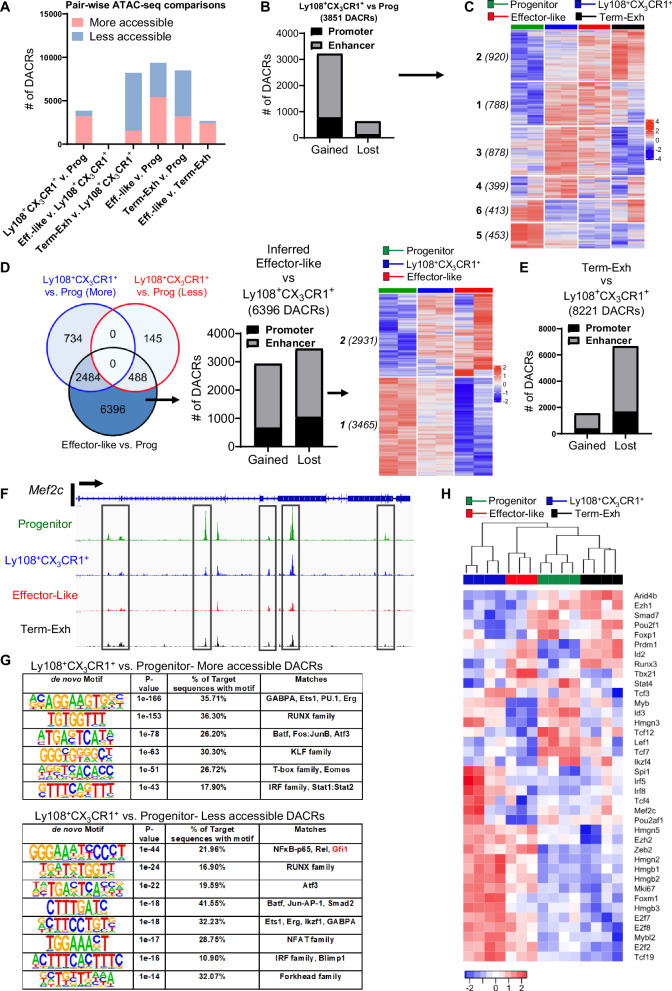
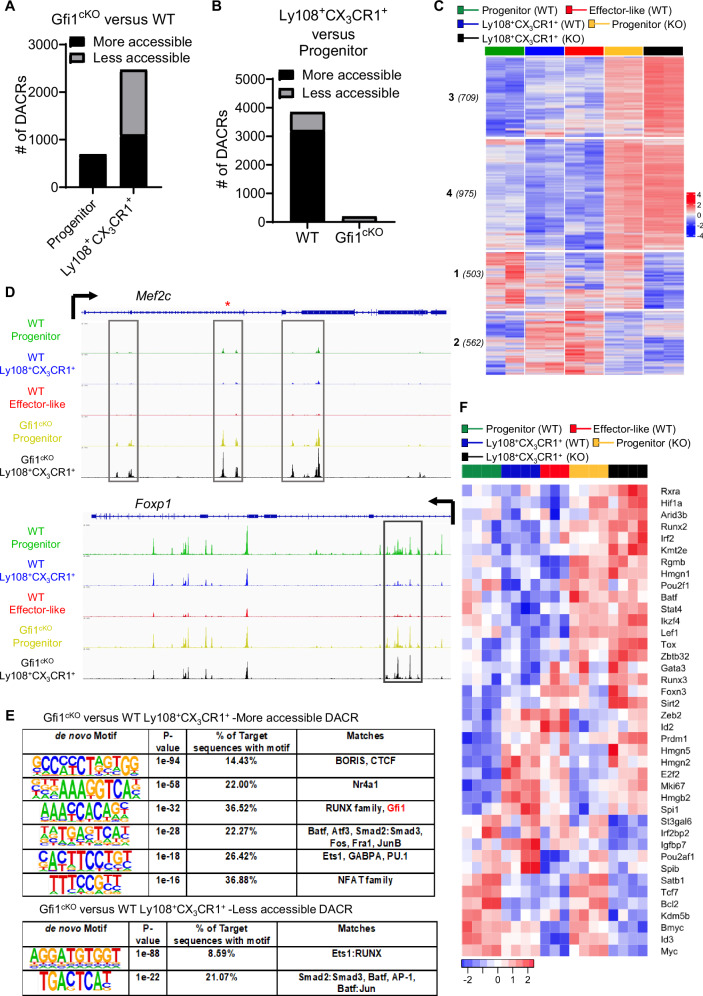
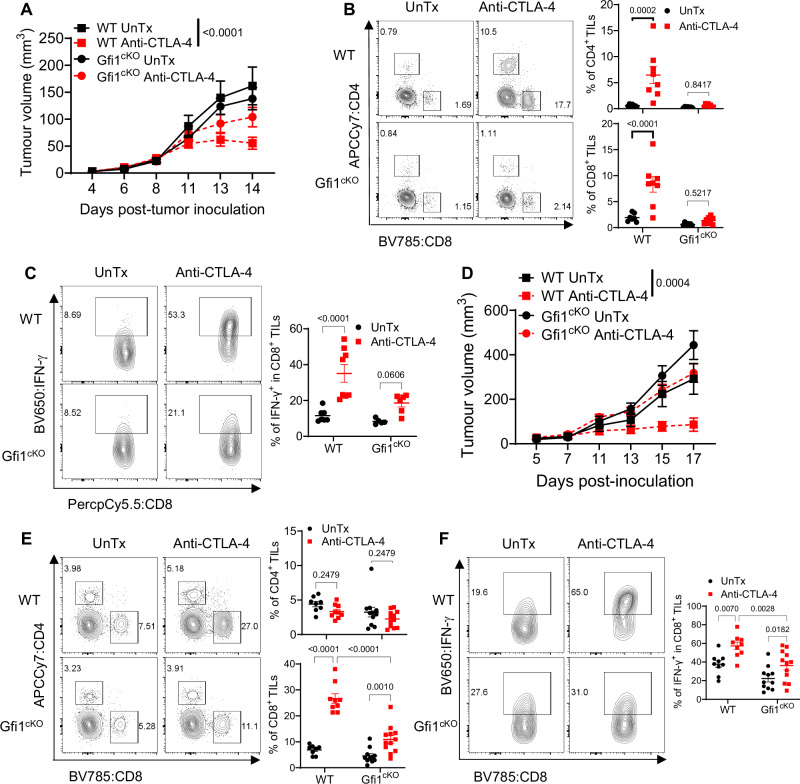
During chronic infection and tumor progression, CD8+ T cells lose their effector functions and become exhausted. These exhausted CD8+ T cells are heterogeneous and comprised of progenitors that give rise to effector-like or terminally-exhausted cells. The precise cues and mechanisms directing subset formation are incompletely understood. Here, we show that growth factor independent-1 (Gfi1) is dynamically regulated in exhausted CD8+ T cells. During chronic LCMV Clone 13 infection, a previously under-described Ly108+CX3CR1+ subset expresses low levels of Gfi1 while other established subsets have high expression. Ly108+CX3CR1+ cells possess distinct chromatin profiles and represent a transitory subset that develops to effector-like and terminally-exhausted cells, a process dependent on Gfi1. Similarly, Gfi1 in tumor-infiltrating CD8+ T cells is required for the formation of terminally differentiated cells and endogenous as well as anti-CTLA-induced anti-tumor responses. Taken together, Gfi1 is a key regulator of the subset formation of exhausted CD8+ T cells.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



Update of
-
Gfi1 controls the formation of effector CD8 T cells during chronic infection and cancer.bioRxiv [Preprint]. 2024 Apr 21:2024.04.18.579535. doi: 10.1101/2024.04.18.579535. bioRxiv. 2024. Update in: Nat Commun. 2025 May 15;16(1):4542. doi: 10.1038/s41467-025-59784-1. PMID: 38659890 Free PMC article. Updated. Preprint.
Similar articles
-
Gfi1 controls the formation of effector CD8 T cells during chronic infection and cancer.bioRxiv [Preprint]. 2024 Apr 21:2024.04.18.579535. doi: 10.1101/2024.04.18.579535. bioRxiv. 2024. Update in: Nat Commun. 2025 May 15;16(1):4542. doi: 10.1038/s41467-025-59784-1. PMID: 38659890 Free PMC article. Updated. Preprint.
-
T Cell Receptor Diversity and Lineage Relationship between Virus-Specific CD8 T Cell Subsets during Chronic Lymphocytic Choriomeningitis Virus Infection.J Virol. 2020 Sep 29;94(20):e00935-20. doi: 10.1128/JVI.00935-20. Print 2020 Sep 29. J Virol. 2020. PMID: 32759317 Free PMC article.
-
BATF regulates progenitor to cytolytic effector CD8+ T cell transition during chronic viral infection.Nat Immunol. 2021 Aug;22(8):996-1007. doi: 10.1038/s41590-021-00965-7. Epub 2021 Jul 19. Nat Immunol. 2021. PMID: 34282329 Free PMC article.
-
GFI1-driven transcriptional and epigenetic programs maintain CD8+ T cell stemness and persistence.Nat Immunol. 2025 Jun;26(6):894-907. doi: 10.1038/s41590-025-02151-5. Epub 2025 May 15. Nat Immunol. 2025. PMID: 40374731 Free PMC article.
-
Exhausted CD8+ T cells face a developmental fork in the road.Trends Immunol. 2023 Apr;44(4):276-286. doi: 10.1016/j.it.2023.02.006. Epub 2023 Mar 11. Trends Immunol. 2023. PMID: 36907685 Free PMC article. Review.
References
-
- Utzschneider, D. T. et al. T Cell factor 1-expressing memory-like CD8(+) T cells sustain the immune response to chronic viral infections. Immunity45, 415–427 (2016). - PubMed
MeSH terms
Substances
Grants and funding
- CRI4342/Cancer Research Institute (CRI)
- 91-022-19/American Cancer Society (American Cancer Society, Inc.)
- R01 CA279849/CA/NCI NIH HHS/United States
- R21 CA259721/CA/NCI NIH HHS/United States
- R01 AI156290/AI/NIAID NIH HHS/United States
- 5R01AI156290/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- V2018-023/V Foundation for Cancer Research (V Foundation)
- ME210108/U.S. Department of Defense (United States Department of Defense)
- 1R01CA279849-01A1, 1R21CA230475-01A1, 1R21CA259721-01A1, 5R01AI156290/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R21 CA230475/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
