

Transcription factors form a ternary complex with NIPBL/MAU2 to localize cohesin at enhancers
- PMID: 40377219
- PMCID: PMC12082454
- DOI: 10.1093/nar/gkaf415
Transcription factors form a ternary complex with NIPBL/MAU2 to localize cohesin at enhancers
Abstract
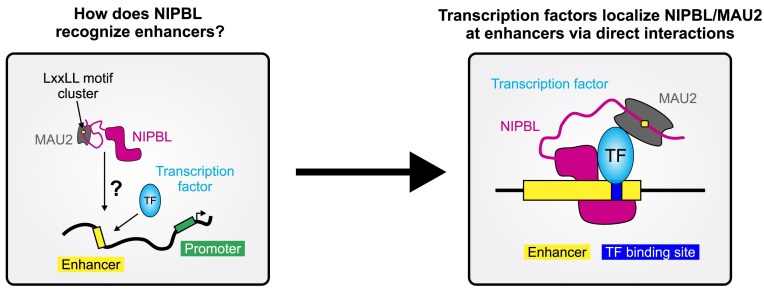
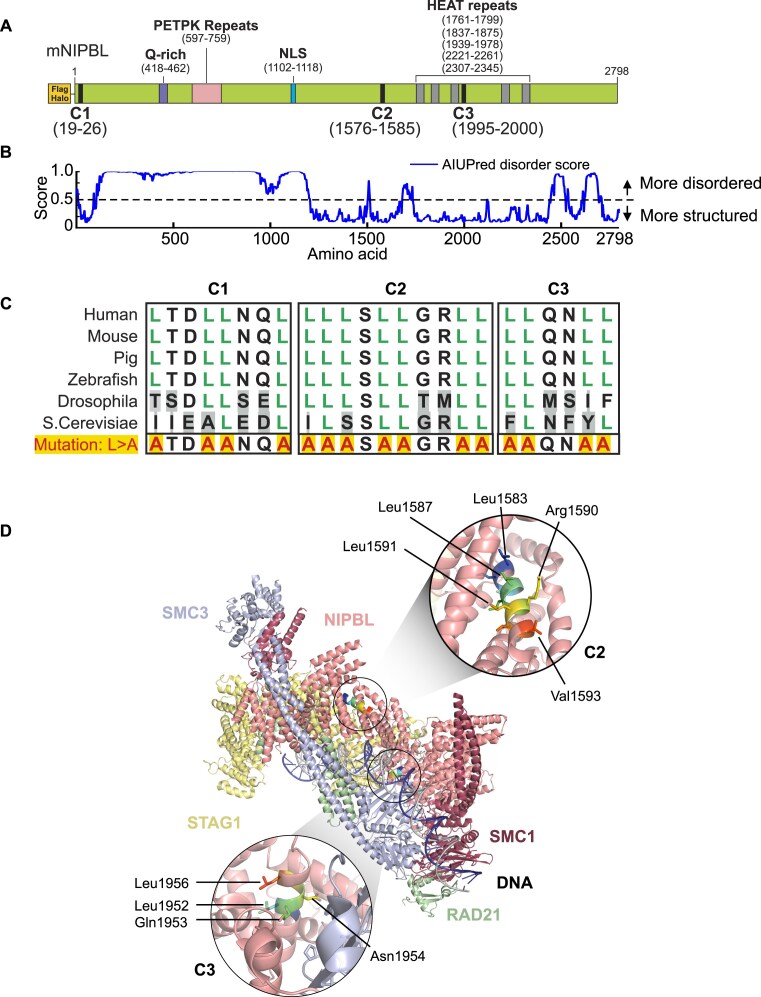
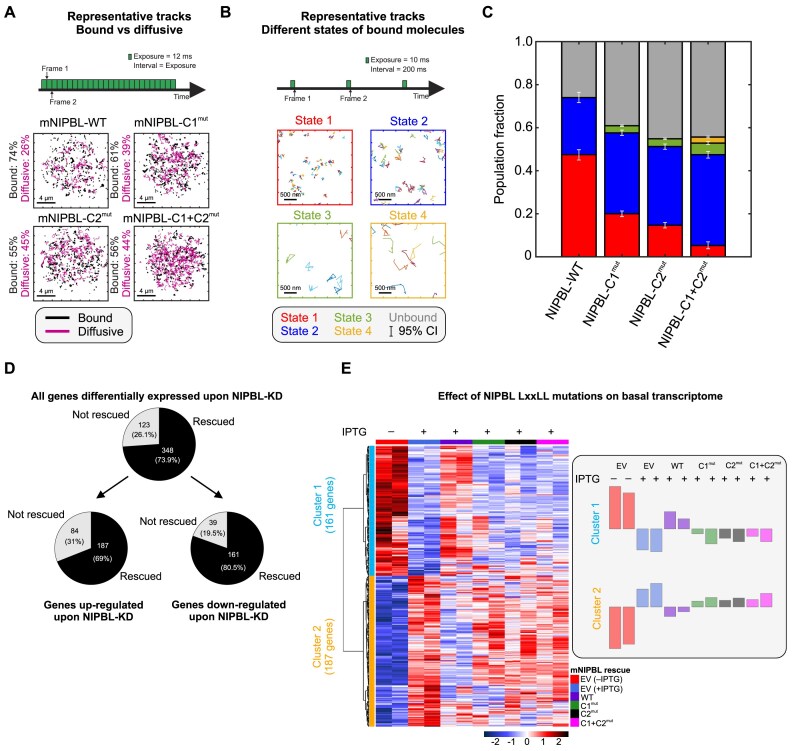
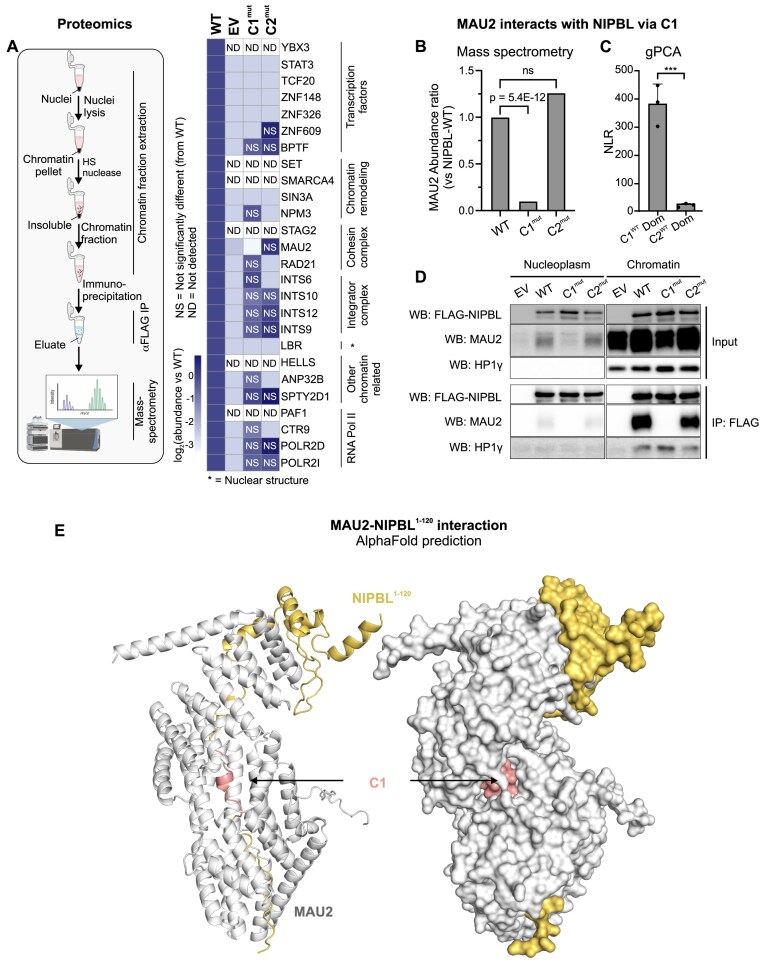
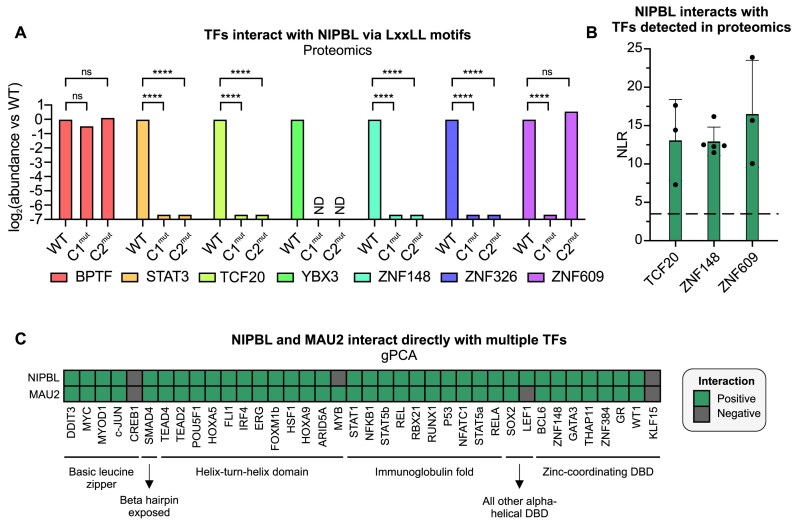
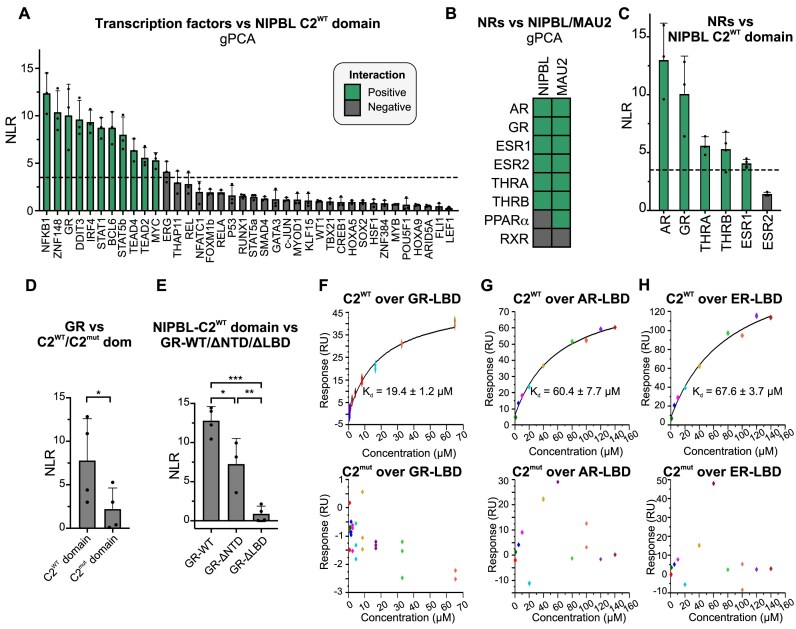
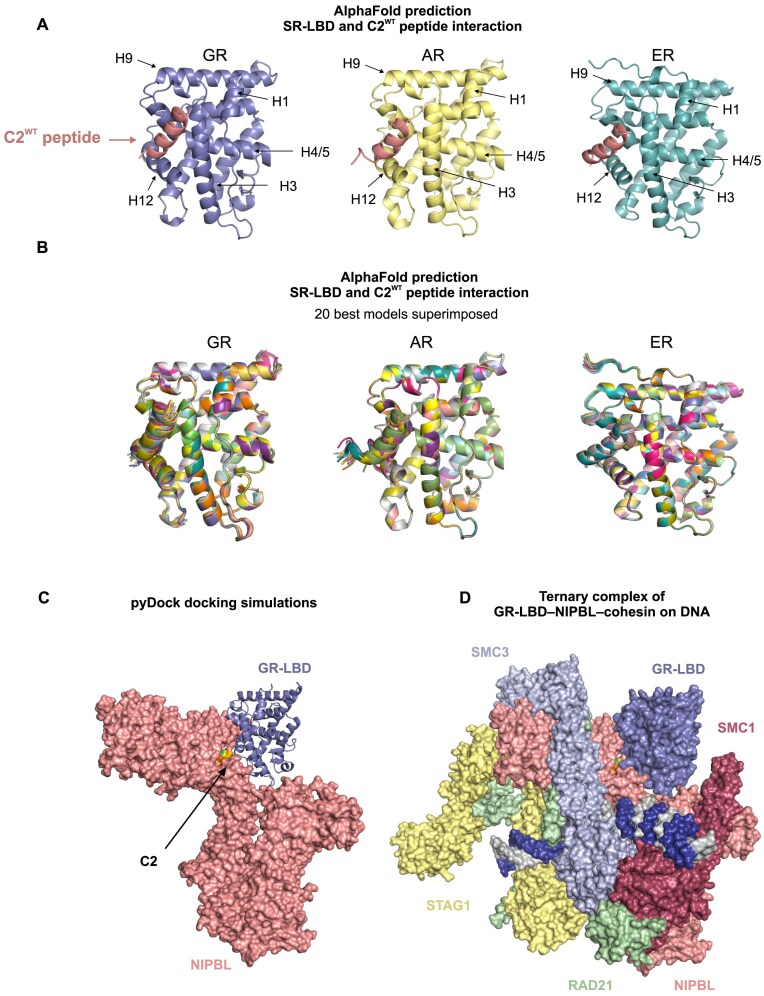
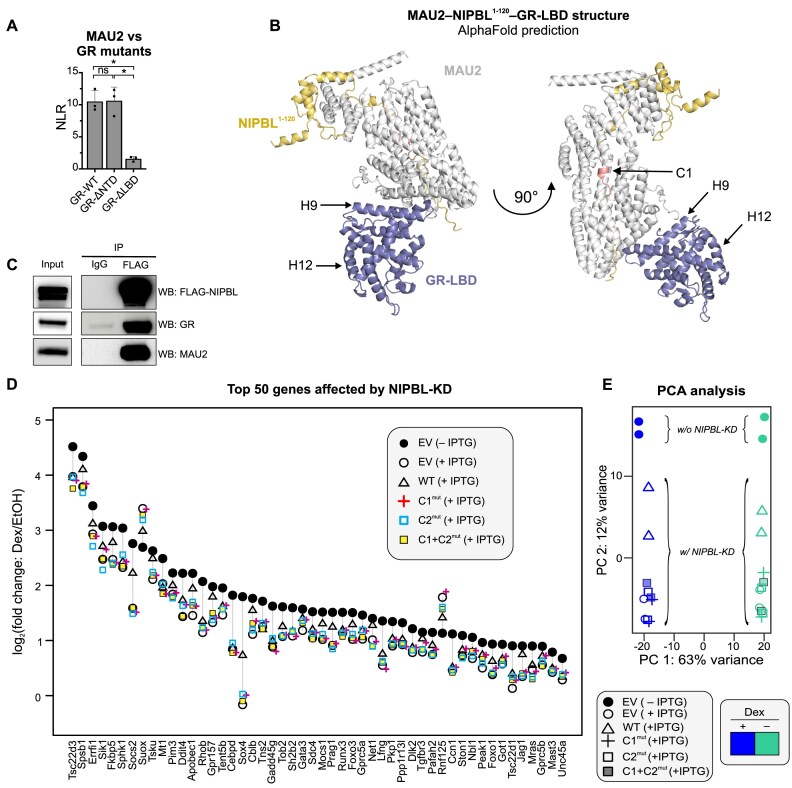
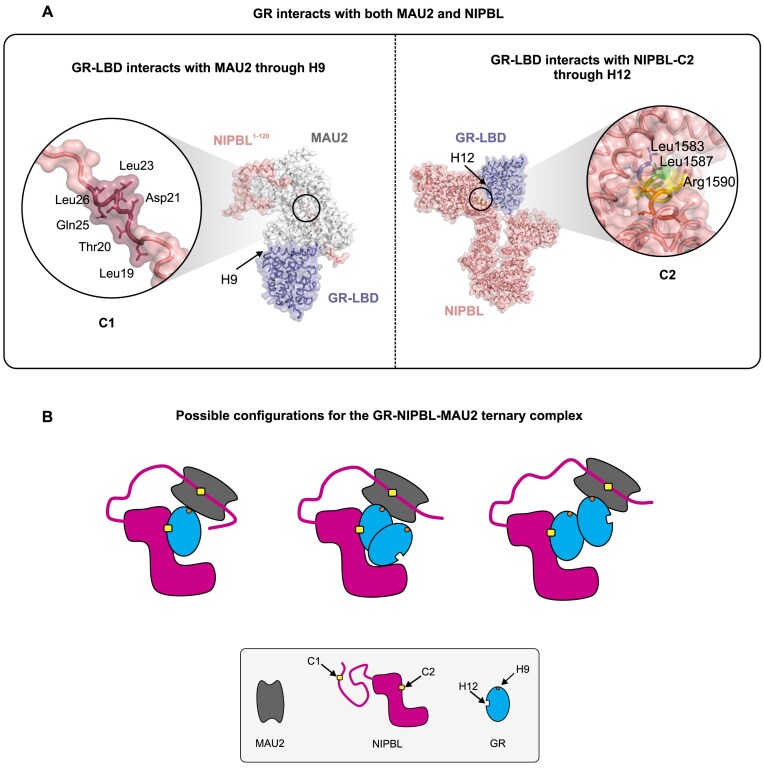
While the cohesin complex is a key player in genome architecture, how it localizes to specific chromatin sites is not understood. Recently, we and others have proposed that direct interactions with transcription factors lead to the localization of the cohesin-loader complex (NIPBL/MAU2) within enhancers. Here, we identify two clusters of LxxLL motifs within the NIPBL sequence that regulate NIPBL dynamics, interactome, and NIPBL-dependent transcriptional programs. One of these clusters interacts with MAU2 and is necessary for the maintenance of the NIPBL-MAU2 heterodimer. The second cluster binds specifically to the ligand-binding domains of steroid receptors. For the glucocorticoid receptor (GR), we examine in detail its interaction surfaces with NIPBL and MAU2. Using AlphaFold2 and molecular docking algorithms, we uncover a GR-NIPBL-MAU2 ternary complex and describe its importance in GR-dependent gene regulation. Finally, we show that multiple transcription factors interact with NIPBL-MAU2, likely using interfaces other than those characterized for GR.
Published by Oxford University Press on behalf of Nucleic Acids Research 2025.
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Transcription factors form a ternary complex with NIPBL/MAU2 to localize cohesin at enhancers.bioRxiv [Preprint]. 2025 Feb 19:2024.12.09.627537. doi: 10.1101/2024.12.09.627537. bioRxiv. 2025. Update in: Nucleic Acids Res. 2025 May 10;53(9):gkaf415. doi: 10.1093/nar/gkaf415. PMID: 39713324 Free PMC article. Updated. Preprint.
Similar articles
-
Transcription factors form a ternary complex with NIPBL/MAU2 to localize cohesin at enhancers.bioRxiv [Preprint]. 2025 Feb 19:2024.12.09.627537. doi: 10.1101/2024.12.09.627537. bioRxiv. 2025. Update in: Nucleic Acids Res. 2025 May 10;53(9):gkaf415. doi: 10.1093/nar/gkaf415. PMID: 39713324 Free PMC article. Updated. Preprint.
-
NIPBL and STAG1 enable loop extrusion by providing differential DNA-cohesin affinity.Proc Natl Acad Sci U S A. 2025 Aug 12;122(32):e2514190122. doi: 10.1073/pnas.2514190122. Epub 2025 Aug 5. Proc Natl Acad Sci U S A. 2025. PMID: 40763028 Free PMC article.
-
Site-to-site mutational dissection of fission yeast cohesin reveals its dynamics.G3 (Bethesda). 2025 Jul 9;15(7):jkaf111. doi: 10.1093/g3journal/jkaf111. G3 (Bethesda). 2025. PMID: 40388360 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Interventions for central serous chorioretinopathy: a network meta-analysis.Cochrane Database Syst Rev. 2025 Jun 16;6(6):CD011841. doi: 10.1002/14651858.CD011841.pub3. Cochrane Database Syst Rev. 2025. PMID: 40522203
Cited by
-
Bile acids target an exposed cavity in the glucocorticoid receptor modulating receptor self-assembly, chromatin binding and transcriptional activity.bioRxiv [Preprint]. 2025 May 16:2025.05.13.653693. doi: 10.1101/2025.05.13.653693. bioRxiv. 2025. PMID: 40463087 Free PMC article. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
