

Dynamics of bacterial operons during genome-wide stresses is influenced by premature terminations and internal promoters
- PMID: 40378216
- PMCID: PMC12083536
- DOI: 10.1126/sciadv.adl3570
Dynamics of bacterial operons during genome-wide stresses is influenced by premature terminations and internal promoters
Abstract
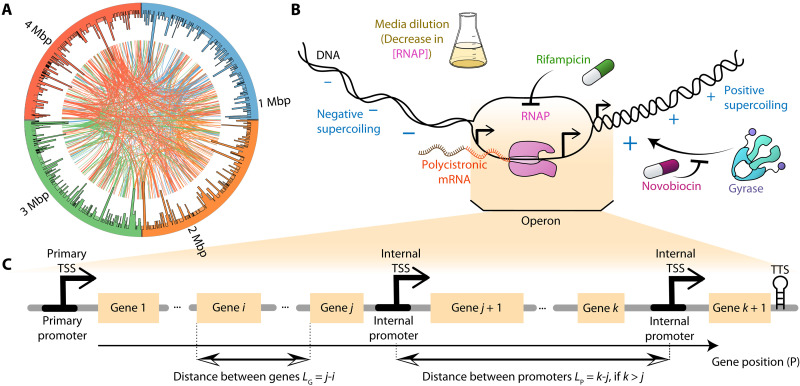
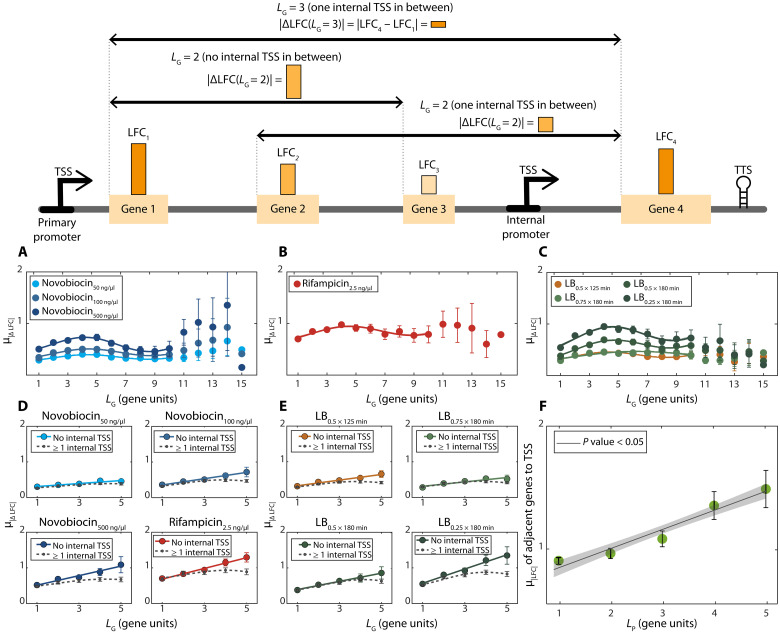
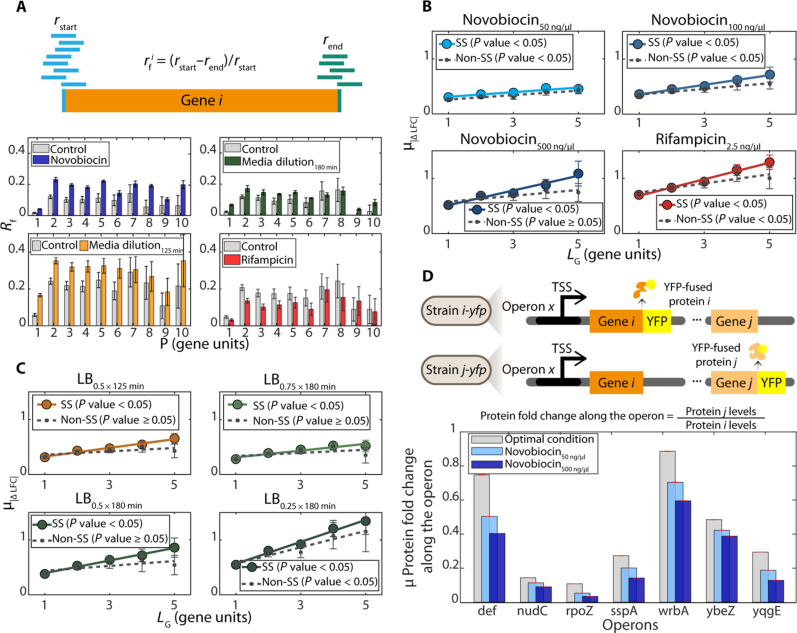
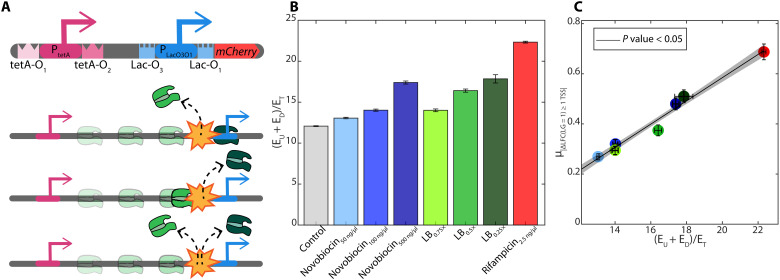
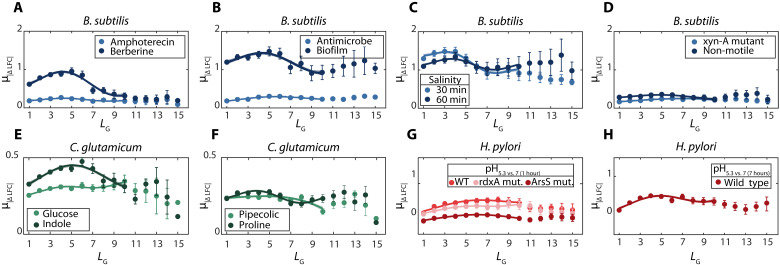
Bacterial gene networks have operons, each coordinating several genes under a primary promoter. Half of the operons in Escherichia coli have been reported to also contain internal promoters. We studied their role during genome-wide stresses targeting key transcription regulators, RNA polymerase (RNAP) and gyrase. Our results suggest that operons' responses are influenced by stress-related changes in premature elongation terminations and internal promoters' activity. Globally, this causes the responses of genes in the same operon to differ with the distance between them in a wave-like pattern. Meanwhile, premature terminations are affected by positive supercoiling buildup, collisions between elongating and promoter-bound RNAPs, and local regulatory elements. We report similar findings in E. coli under other stresses and in evolutionarily distant bacteria Bacillus subtilis, Corynebacterium glutamicum, and Helicobacter pylori. Our results suggest that the strength, number, and positioning of operons' internal promoters might have evolved to compensate for premature terminations, providing distal genes similar response strengths.
Figures
Similar articles
-
Inferring regulatory elements from a whole genome. An analysis of Helicobacter pylori sigma(80) family of promoter signals.J Mol Biol. 2000 Mar 24;297(2):335-53. doi: 10.1006/jmbi.2000.3576. J Mol Biol. 2000. PMID: 10715205
-
Transcription Attenuation in Synthetic Promoters in Nonoverlapping Tandem Formation.Biochemistry. 2024 Aug 20;63(16):2009-2022. doi: 10.1021/acs.biochem.4c00012. Epub 2024 Jul 12. Biochemistry. 2024. PMID: 38997112 Free PMC article.
-
Conserved Units of Co-Expression in Bacterial Genomes: An Evolutionary Insight into Transcriptional Regulation.PLoS One. 2016 May 19;11(5):e0155740. doi: 10.1371/journal.pone.0155740. eCollection 2016. PLoS One. 2016. PMID: 27195891 Free PMC article.
-
The effects of DNA supercoiling on the expression of operons of the ilv regulon of Escherichia coli suggest a physiological rationale for divergently transcribed operons.Mol Microbiol. 2001 Mar;39(5):1109-15. doi: 10.1111/j.1365-2958.2001.02309.x. Mol Microbiol. 2001. PMID: 11251829 Review.
-
Sigma factors and promoters in Corynebacterium glutamicum.J Biotechnol. 2011 Jul 10;154(2-3):101-13. doi: 10.1016/j.jbiotec.2011.01.017. Epub 2011 Jan 26. J Biotechnol. 2011. PMID: 21277915 Review.
References
-
- Jacob F., Monod J., On the regulation of gene activity. Cold Spring Harb. Symp. Quant. Biol. 26, 193–211 (1961). - PubMed
-
- Browning D. F., Busby S. J. W., The regulation of bacterial transcription initiation. Nat. Rev. Microbiol. 2, 57–65 (2004). - PubMed
-
- McClure W. R., Mechanism and control of transcription initiation in prokaryotes. Annu. Rev. Biochem. 54, 171–204 (1985). - PubMed
-
- Swain P. S., Efficient attenuation of stochasticity in gene expression through post-transcriptional control. J. Mol. Biol. 344, 965–976 (2004). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
