

CD44 is a macrophage receptor for TcdB from Clostridioides difficile that via its lysine-158 succinylation contributes to inflammation
- PMID: 40383912
- PMCID: PMC12091907
- DOI: 10.1080/19490976.2025.2506192
CD44 is a macrophage receptor for TcdB from Clostridioides difficile that via its lysine-158 succinylation contributes to inflammation
Erratum in
-
Correction.Gut Microbes. 2025 Dec;17(1):2532985. doi: 10.1080/19490976.2025.2532985. Epub 2025 Jul 11. Gut Microbes. 2025. PMID: 40650384 Free PMC article. No abstract available.
Abstract
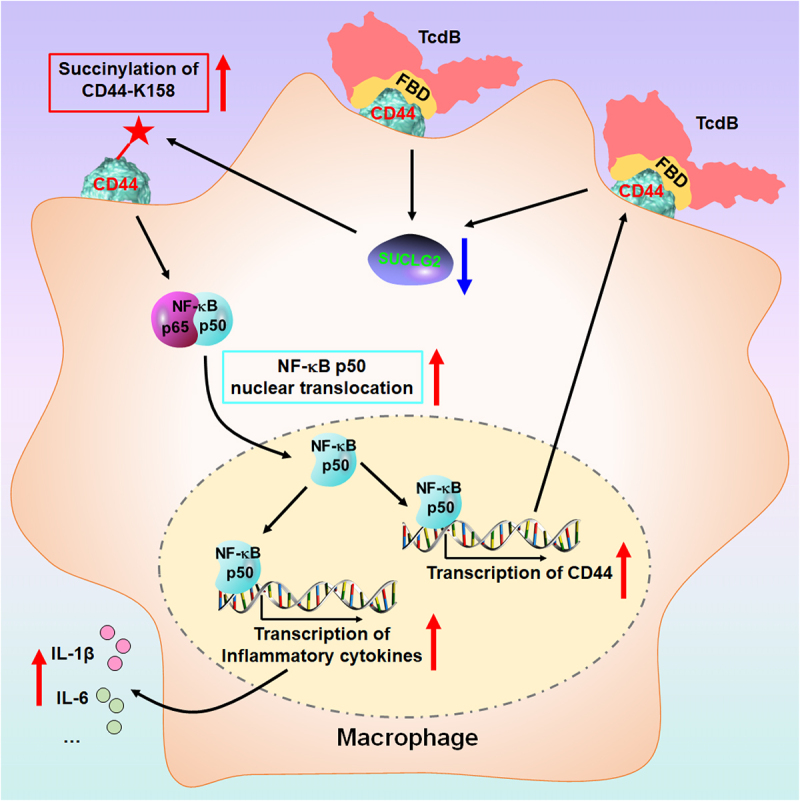
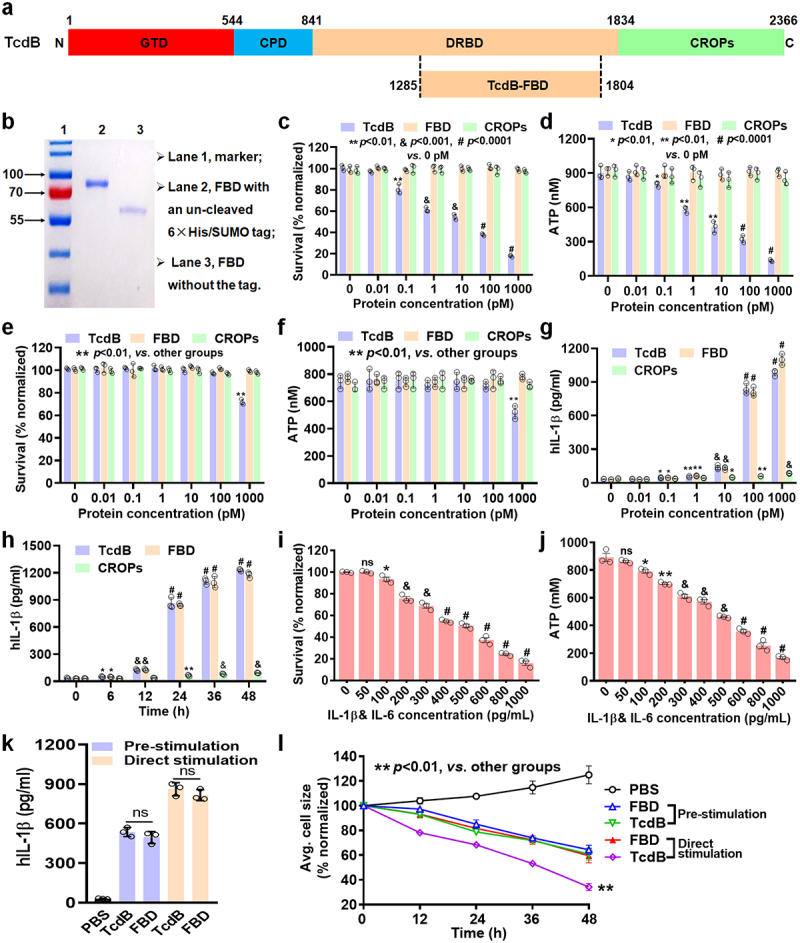
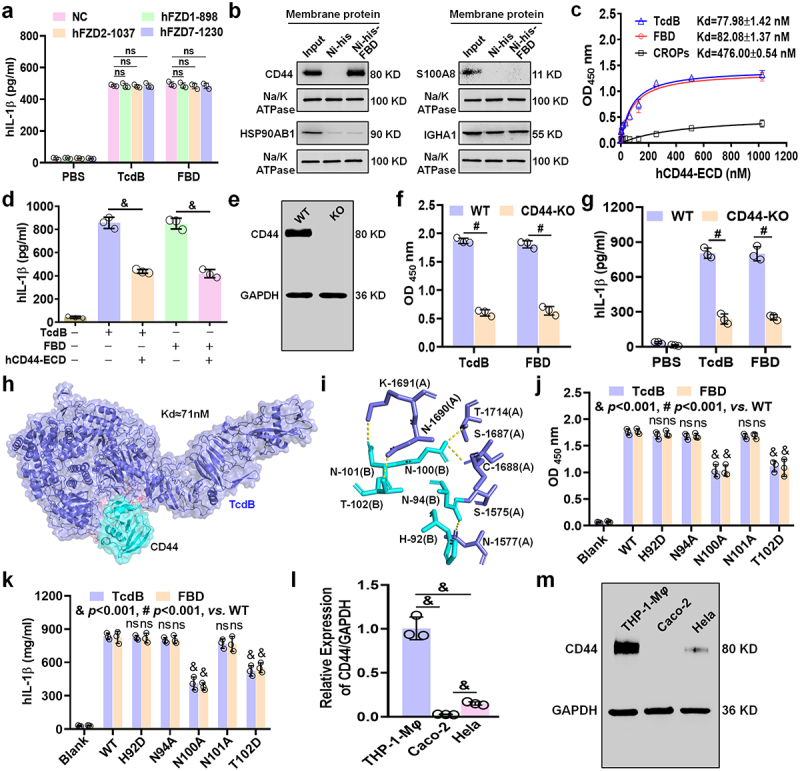
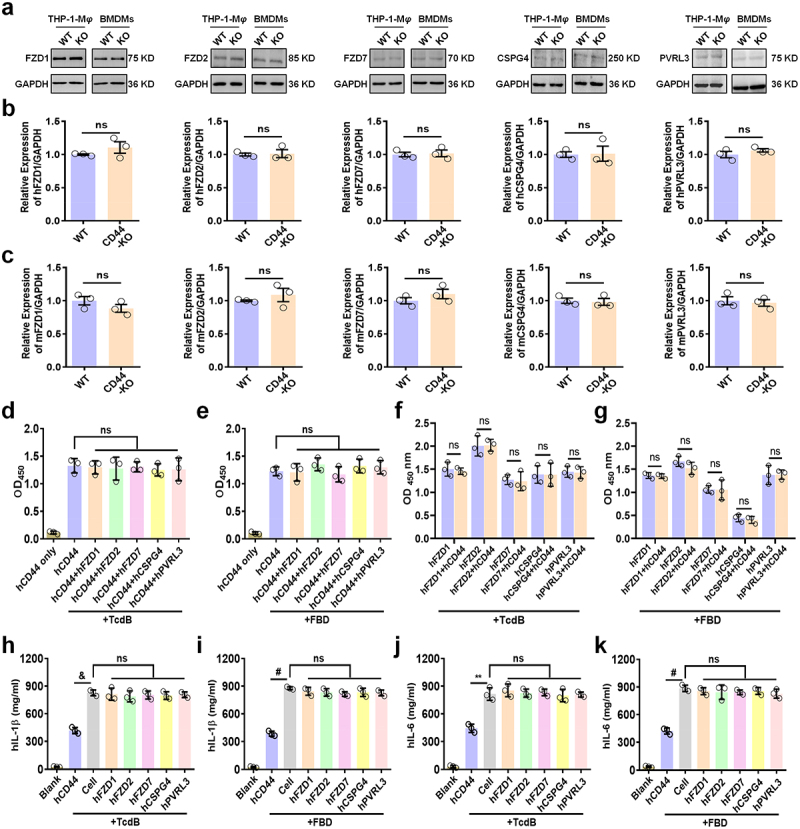
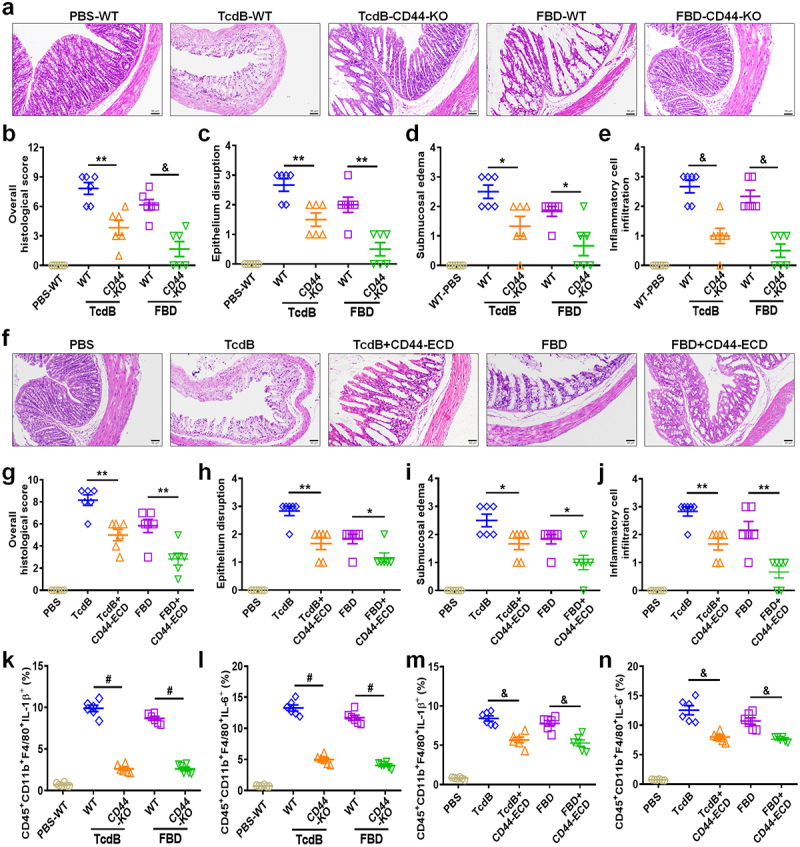
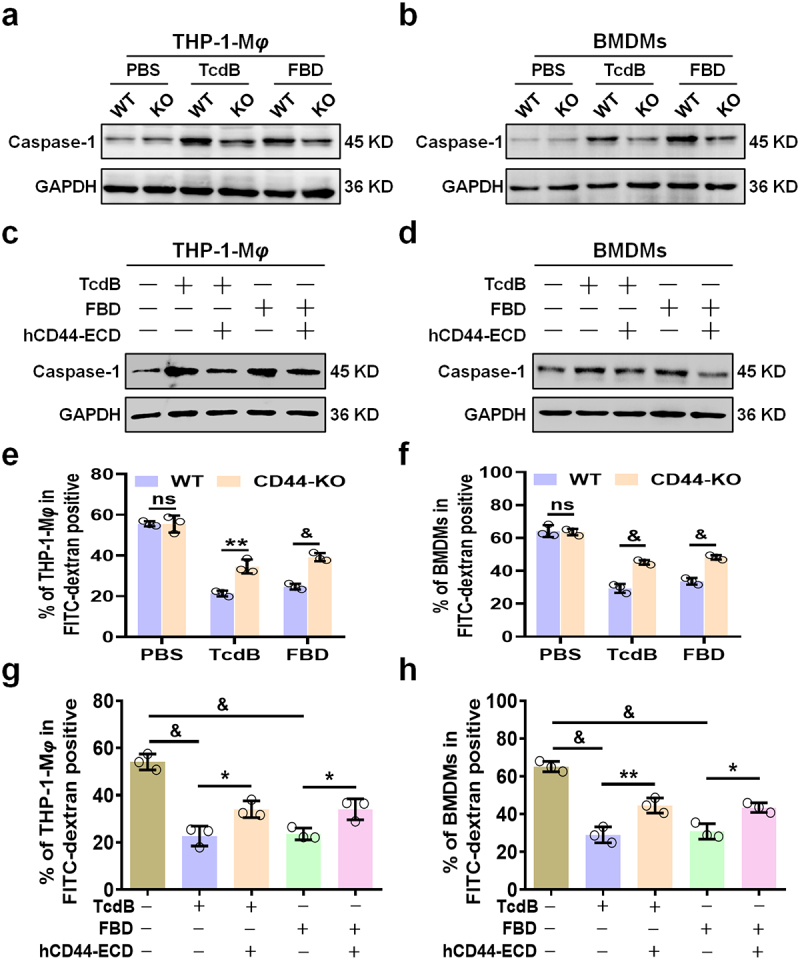
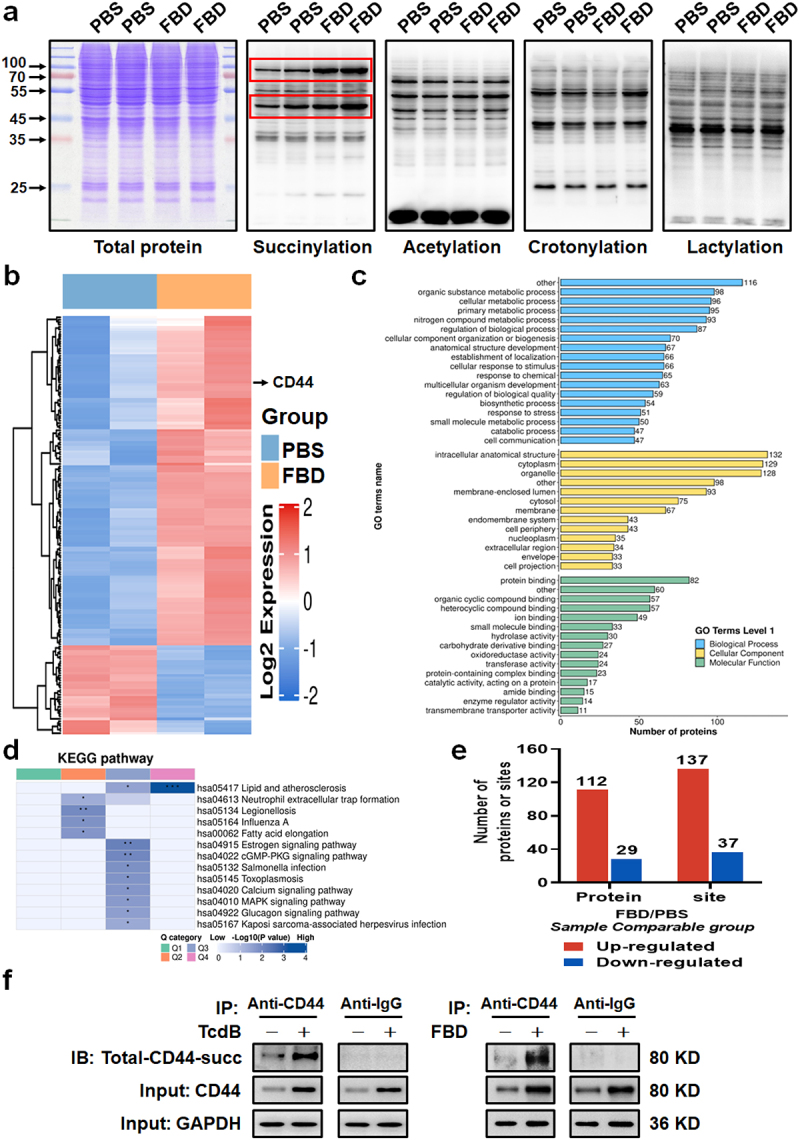
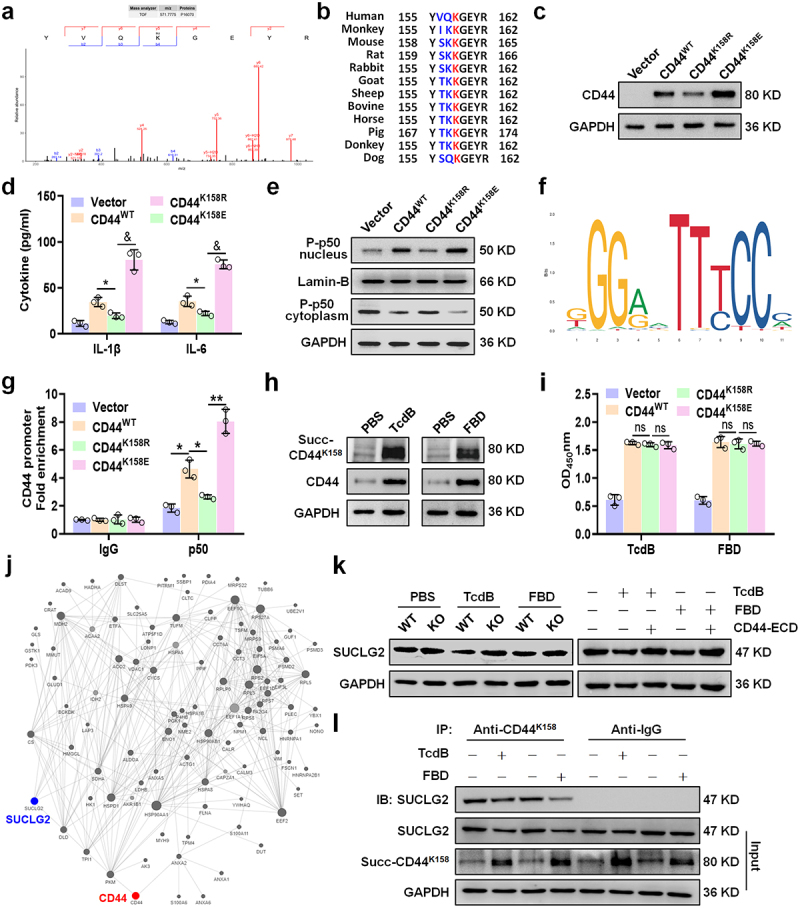
Toxin B (TcdB) is a critical virulence factor in Clostridioides difficile-associated disease (CDAD), which activates macrophages to promote inflammation and epithelial damage. However, the mechanism by which TcdB targets inflammation-related receptors on the macrophage surface and the underlying molecular mechanisms remain unknown. The frizzled-binding domain of TcdB (TcdB-FBD) is a promising target of TcdB. Here, FBD was found to trigger macrophage inflammation, similar to TcdB, but did not induce cytotoxicity. Thus, using FBD as a bait protein, macrophage CD44 was identified as an inflammation-related receptor for TcdB/FBD. The role of CD44 was confirmed by CRISPR/Cas9-mediated gene knockout in macrophages and CD44 knockout mice. Using 4-D label-free succinylation quantitative modification proteomics, we demonstrated that TcdB/FBD binds to CD44 in macrophages, promotes CD44 K158 succinylation via SUCLG2 suppression, and enhances NF-κB translocation/transcriptional activity, thereby driving inflammation. Finally, blocking the binding of TcdB to CD44 was demonstrated as a favorable strategy for inhibiting TcdB-mediated macrophage inflammation. This study not only provides a new therapeutic target for the prevention and treatment of CDAD but also elucidates a new molecular mechanism underlying the inflammatory effect of TcdB via the TcdB/FBD-CD44 axis.
Keywords: CD44; TcdB; inflammation; receptor; succinylation.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
References
-
- Di Bella S, Sanson G, Monticelli J, Zerbato V, Principe L, Giuffre M, Pipitone G, Luzzati R, Staley C.. Clostridioides difficile infection: history, epidemiology, risk factors, prevention, clinical manifestations, treatment, and future options. Clin Microbiol Rev. 2024;37(2):e0013523. doi: 10.1128/cmr.00135-23. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous