

This is a preprint.
RPE65 variant p.(E519K) causes a novel dominant adult-onset maculopathy in 83 affected individuals
- PMID: 40386434
- PMCID: PMC12083654
- DOI: 10.21203/rs.3.rs-5849564/v2
RPE65 variant p.(E519K) causes a novel dominant adult-onset maculopathy in 83 affected individuals
Update in
-
RPE65 Variant p.(E519K) Causes a Novel Dominant Adult-Onset Maculopathy in 83 Affected Individuals.Invest Ophthalmol Vis Sci. 2025 Sep 2;66(12):53. doi: 10.1167/iovs.66.12.53. Invest Ophthalmol Vis Sci. 2025. PMID: 40985799 Free PMC article.
Abstract
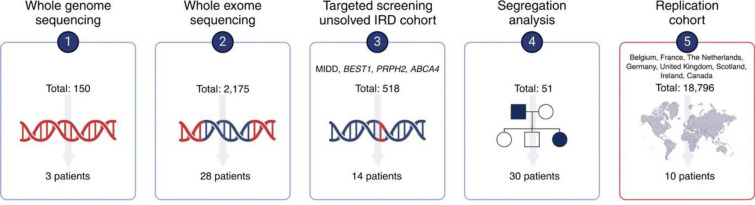
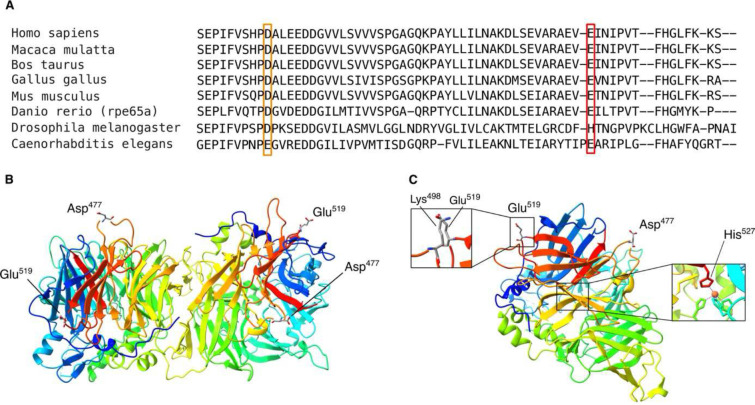
Recessive RPE65-related retinopathy is an inherited retinal disease (IRD) that is a well-established target for gene therapy. Dominant RPE65-related retinopathy, however, due to Irish founder variant p.(D477G), is extremely rare. Here, we report the discovery, replication and characterization of a novel dominant retinopathy caused by RPE65 variant p.(E519K), identified in 83 individuals of European ancestry across IRD registries (Belgian discovery cohort, n=2,873; replication cohort, n=18,796). Long-read sequencing-based haplotyping revealed a shared region of 464 kb, supporting a founder effect. Genotype-phenotype data support dominant inheritance and phenotypic variability respectively, characterized by late-onset macular dystrophy with two main subtypes, a pathognomonic mottled subtype and a pattern dystrophy subtype. Functional studies showed that the p.(E519K) variant affects RPE65 enzymatic activity, correlating with lower protein expression. Protein modelling and cellular thermal shift assays further supported a destabilizing effect on protein structure. Overall, our work provides strong genetic, clinical, molecular and functional evidence for a novel dominant RPE65 retinopathy in multiple families in Europe and North America due to a Belgian founder variant. This discovery reduces the diagnostic gap in dominant IRD, particularly in individuals of European ancestry. Finally, it lays the foundation for developing therapeutic strategies targeting dominant RPE65 retinopathy.
Keywords: Autosomal dominant; European ancestry; Founder; Inherited Retinal Disease (IRD); RPE65.
Conflict of interest statement
Conflicts of Interests The authors declare no competing interests
Figures



References
-
- Bourne RRA, Steinmetz JD, Saylan M, et al. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: The Right to Sight: An analysis for the Global Burden of Disease Study. Lancet Glob Health. 2021;9(2):e144–e160. doi: 10.1016/S2214-109X(20)30489-7 - DOI - PMC - PubMed
-
- WHO. Blindness and vision impairment. August 10, 2023. Accessed October 1, 2023. https://www.who.int/news-room/fact-sheets/detail/blindness-and-visual-im...
-
- Moiseyev G, Chen Y, Takahashi Y, Wu BX, Ma JX. RPE65 Is the Isomerohydrolase in the Retinoid Visual Cycle.; 2005. www.pnas.orgcgidoi10.1073pnas.0503460102 - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
