

FUT8 Is a Critical Driver of Prostate Tumour Growth and Can Be Targeted Using Fucosylation Inhibitors
- PMID: 40387385
- PMCID: PMC12086987
- DOI: 10.1002/cam4.70959
FUT8 Is a Critical Driver of Prostate Tumour Growth and Can Be Targeted Using Fucosylation Inhibitors
Abstract
Background: An unmet clinical need requires the discovery of new treatments for men facing advanced prostate cancer. Aberrant glycosylation is a universal feature of cancer cells and plays a key role in tumour growth, immune evasion and metastasis. Alterations in tumour glycosylation are closely associated with prostate cancer progression, making glycans promising therapeutic targets. Fucosyltransferase 8 (FUT8) drives core fucosylation by adding α1,6-fucose to the innermost GlcNAc residue on N-glycans. While FUT8 is recognised as a crucial factor in cancer progression, its role in prostate cancer remains poorly understood.
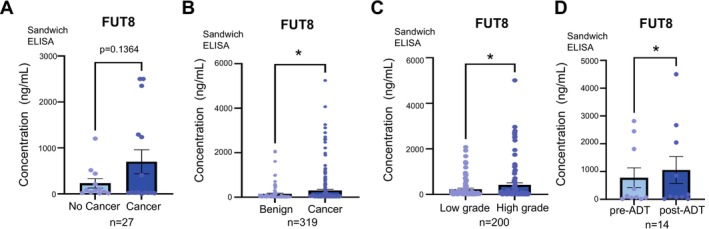
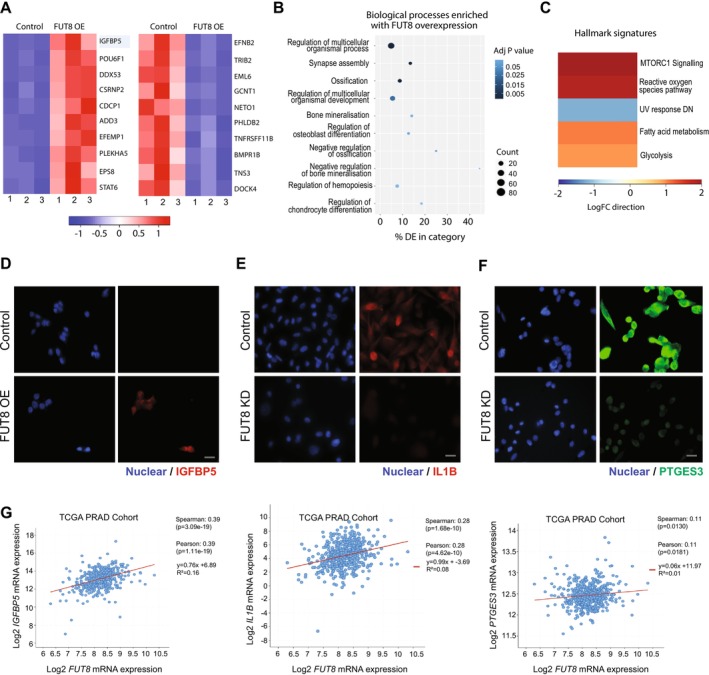
Methods & results: Here, we demonstrate using multiple independent clinical cohorts that FUT8 is upregulated in high grade and metastatic prostate tumours, and in the blood of prostate cancer patients with aggressive disease. Using novel tools, including PhosL lectin immunofluorescence and N-glycan MALDI mass spectrometry imaging (MALDI-MSI), we find FUT8 underpins the biosynthesis of malignant core fucosylated N-glycans in prostate cancer cells and using both in vitro and in vivo models, we find FUT8 promotes prostate tumour growth, cell motility and invasion. Mechanistically we show FUT8 regulates the expression of genes and signalling pathways linked to prostate cancer progression. Furthermore, we find that fucosylation inhibitors can inhibit the activity of FUT8 in prostate cancer to suppress the growth of prostate tumours.
Conclusions: Our study cements FUT8-mediated core fucosylation as an important driver of prostate cancer progression and suggests targeting FUT8 activity for prostate cancer therapy as an exciting area to explore.
Keywords: core fucosylation; fucosylation inhibitors; fucosyltransferase 8 (FUT8); glycans; prostate cancer; therapeutics; tumour growth.
© 2025 The Author(s). Cancer Medicine published by John Wiley & Sons Ltd.
Conflict of interest statement
J.M. and E.S. are shareholders of GlycoScoreDx Ltd. and have filed patents related to this work (GB Patent GB2,594,103 and US Patent App. 17/780,508). J.F.A.P. and E.R. are shareholders of and employed by GlycoTherapeutics B.V. T.J.B. is a shareholder of and scientific advisor of GlycoTherapeutics B.V.; J.F.A.P. and T.J.B. are shareholders of Synvenio B.V. Radboud University and Radboudumc have filed patent applications related to Fucotrim I and Fucotrim II. All other authors declare no conflicts of interest.
Figures




References
-
- Sung H., Ferlay J., Siegel R. L., et al., “Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries,” CA: A Cancer Journal for Clinicians 71, no. 3 (2021): 209–249. - PubMed
-
- Bray F., Laversanne M., Sung H., et al., “Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries,” CA: A Cancer Journal for Clinicians 74, no. 3 (2024): 229–263. - PubMed
-
- Nuhn P., De Bono J. S., Fizazi K., et al., “Update on Systemic Prostate Cancer Therapies: Management of Metastatic Castration‐Resistant Prostate Cancer in the Era of Precision Oncology,” European Urology 75, no. 1 (2019): 88–99. - PubMed
-
- Mateo J., Fizazi K., Gillessen S., et al., “Managing Nonmetastatic Castration‐Resistant Prostate Cancer,” European Urology 75, no. 2 (2019): 285–293. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
