

Uncovering biomarkers for chronic toxoplasmosis detection highlights alternative pathways shaping parasite dormancy
- PMID: 40389644
- PMCID: PMC12254245
- DOI: 10.1038/s44321-025-00252-0
Uncovering biomarkers for chronic toxoplasmosis detection highlights alternative pathways shaping parasite dormancy
Abstract
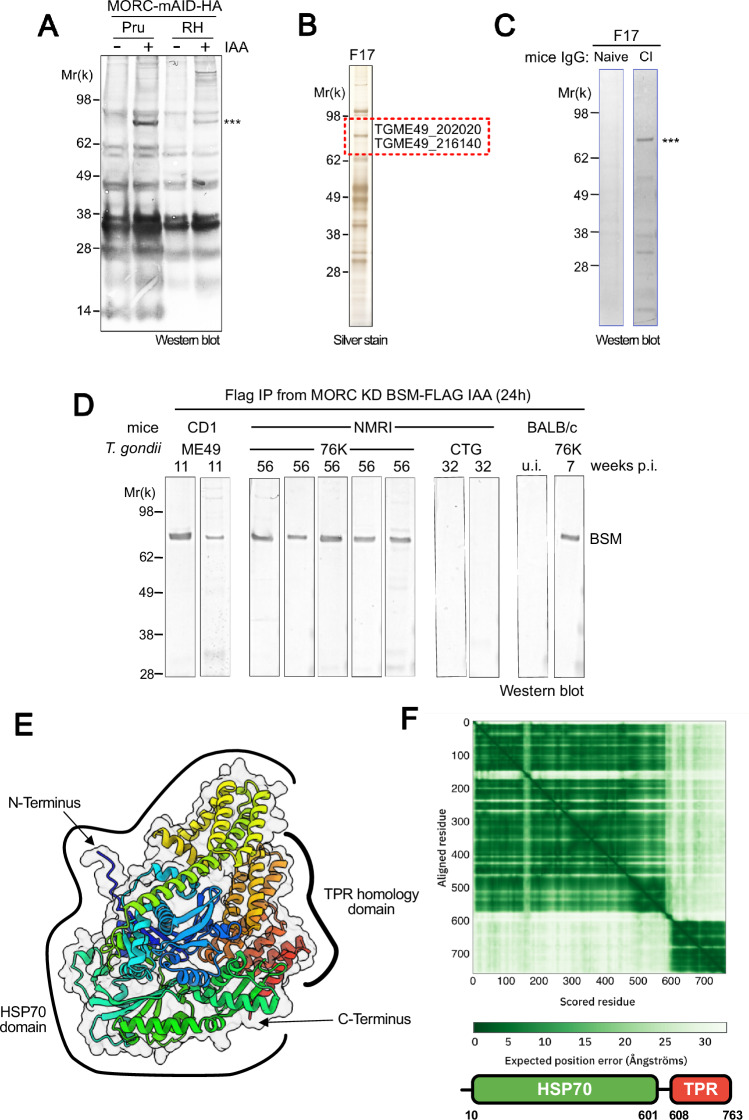
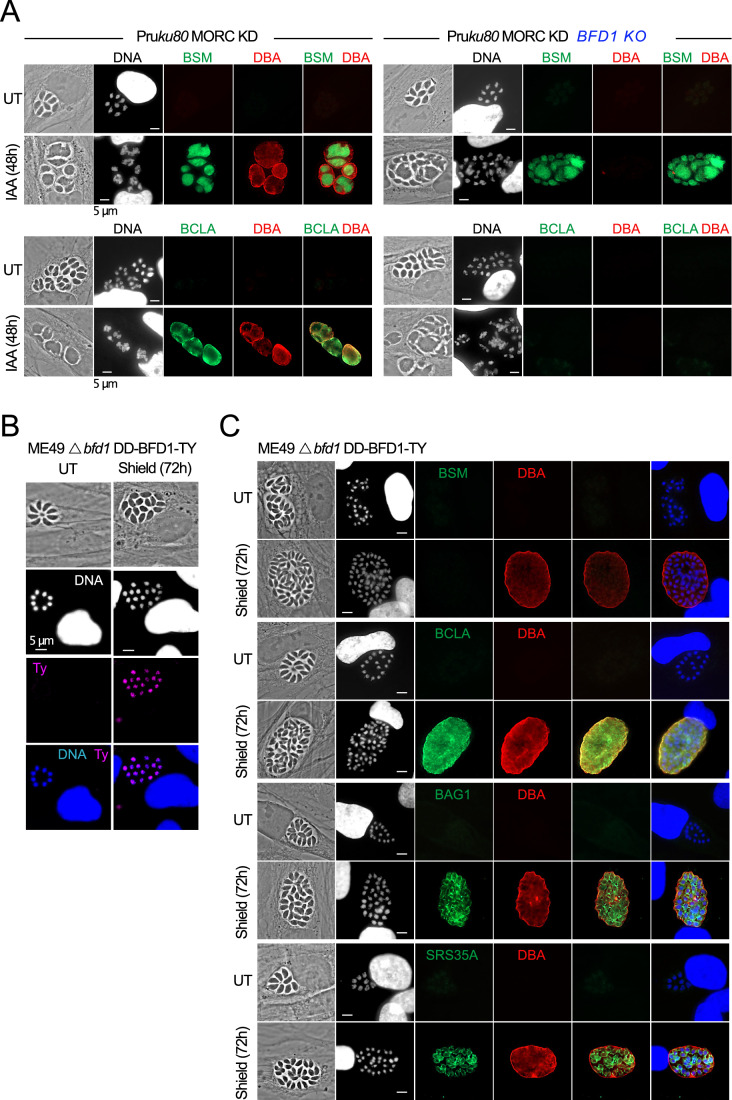
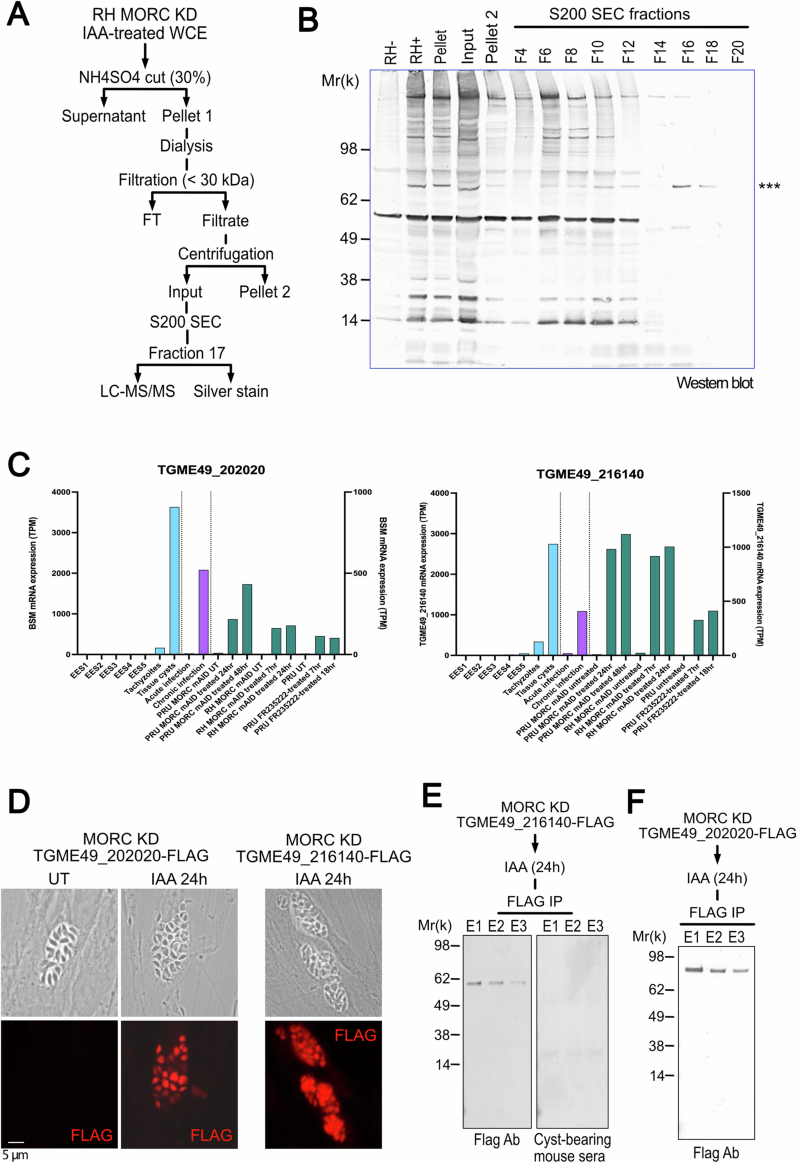
Toxoplasma gondii, a neurotropic protozoan, causes toxoplasmosis, a prevalent zoonotic and food-borne infection, posing significant risks to immunocompromised individuals and congenital cases. The chronic phase, characterized by dormant, cyst-forming bradyzoites, is central to disease progression but is poorly understood due to the lack of serological tests to detect bradyzoite-specific antigens. This study identifies the bradyzoite serological marker (BSM) and cyst-associated BCLA as effective biomarkers for chronic toxoplasmosis. These markers showed high sensitivity and specificity in detecting cyst-bearing mice and had a positivity rate of 30% in humans with prior immunity. Bradyzoite serology helps to discriminate between recent and past infections, with BCLA improving the accuracy of the diagnosis of congenital infections. Mechanistic analyses show that the chromatin modifiers MORC and HDAC3 epistatically regulate BFD1, a key bradyzoite regulator. While BFD1 controls the expression of bradyzoite genes such as BCLA, a specific subset, including BSM, is regulated independently of BFD1. This multilayered regulation complicates the understanding of parasite persistence in humans, but offers promise for improved serologic diagnosis during pregnancy, but also in individuals with mental illness.
Keywords: Toxoplasma gondii; Bradyzoite; Epigenetics; Serology; Transcription.
© 2025. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. MAH, CD, CS, HP, and MPBP are co-inventors of patent PCT/EP2020/081638, covering the use of BCLA as a biomarker for diagnosing chronic toxoplasmosis in humans. MAH, MGR, CS, and MPBP are also co-inventors of a related patent application on using BSM for the same purpose (patent EP24307283.2). Inserm and its subsidiary, Inserm-Transfert, are involved in the patent application. The remaining authors declare no competing interests.
Figures






References
-
- Ben-Harari RR, Connolly MP (2019) High burden and low awareness of toxoplasmosis in the United States. Postgrad Med 131:103–108 - PubMed
-
- Bougdour A, Durandau E, Brenier-Pinchart MP, Ortet P, Barakat M, Kieffer S, Curt-Varesano A, Curt-Bertini RL, Bastien O, Coute Y, Pelloux H, Hakimi MA (2013) Host cell subversion by Toxoplasma GRA16, an exported dense granule protein that targets the host cell nucleus and alters gene expression. Cell Host Microbe 13:489–500 - PubMed
MeSH terms
Substances
Grants and funding
- Project LatentToxoDiag,DS-2022-0017/MSD Avenir
- Project Chairs of Excellence in Biology/Health,Project ToxoNeoSex,ANR-24-CHBS-0008/Agence Nationale de la Recherche (ANR)
- Project ApiMORCing,ANR-21-CE15-0002-01/Agence Nationale de la Recherche (ANR)
- Project ApiNewDrug,ANR-21-CE35-0010-01/Agence Nationale de la Recherche (ANR)
- Laboratoire d'Excellence (LabEx) ParaFrap [ANR-11-LABX-0024]/Agence Nationale de la Recherche (ANR)
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
