

Deciphering the role of histone modifications in memory and exhausted CD8 T cells
- PMID: 40389726
- PMCID: PMC12089470
- DOI: 10.1038/s41598-025-99804-0
Deciphering the role of histone modifications in memory and exhausted CD8 T cells
Abstract
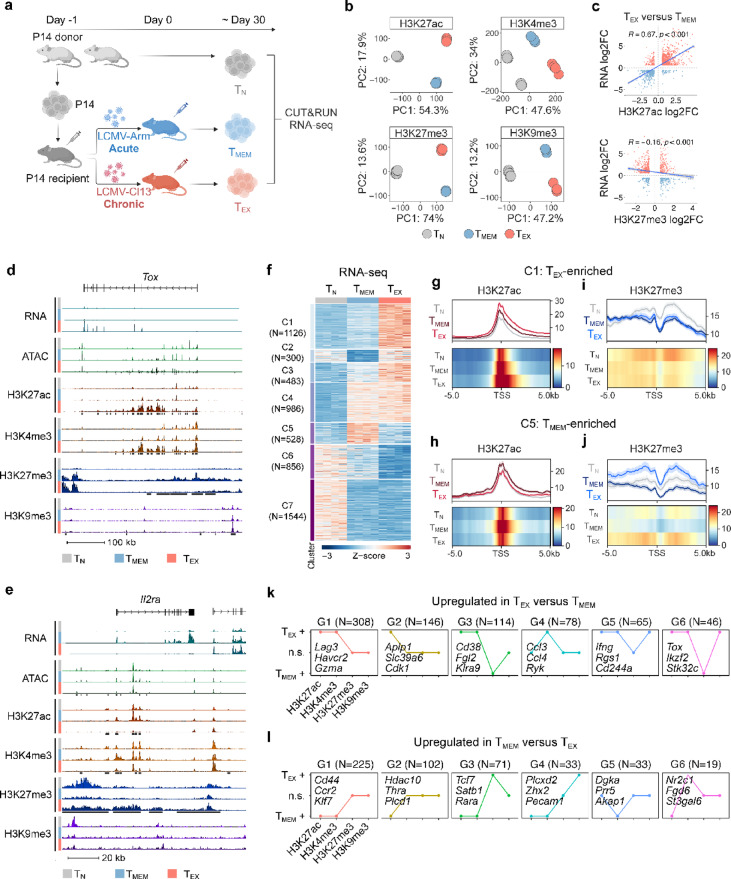
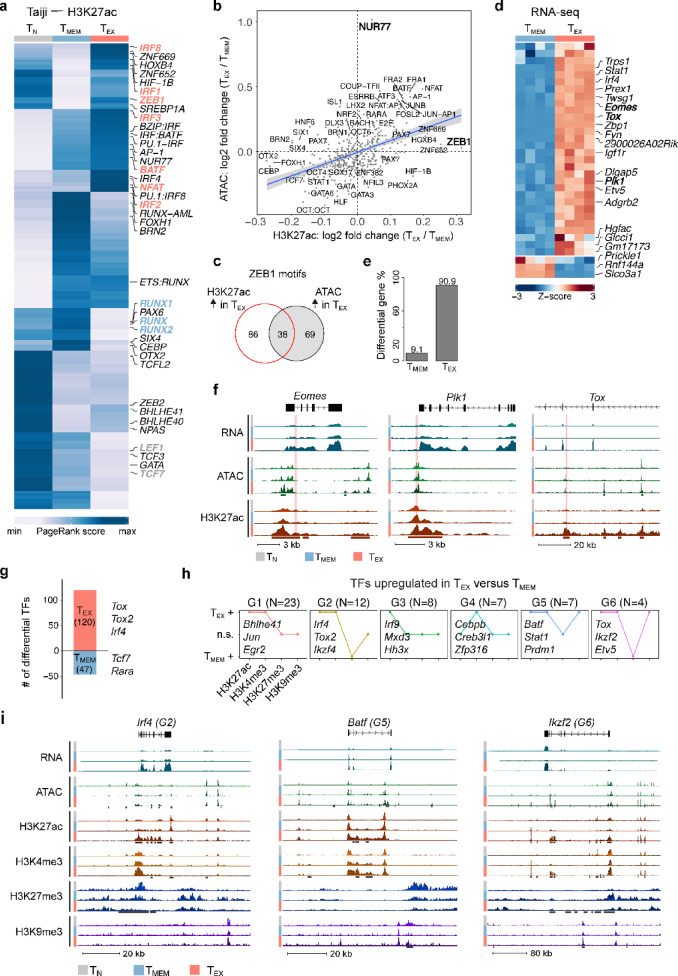
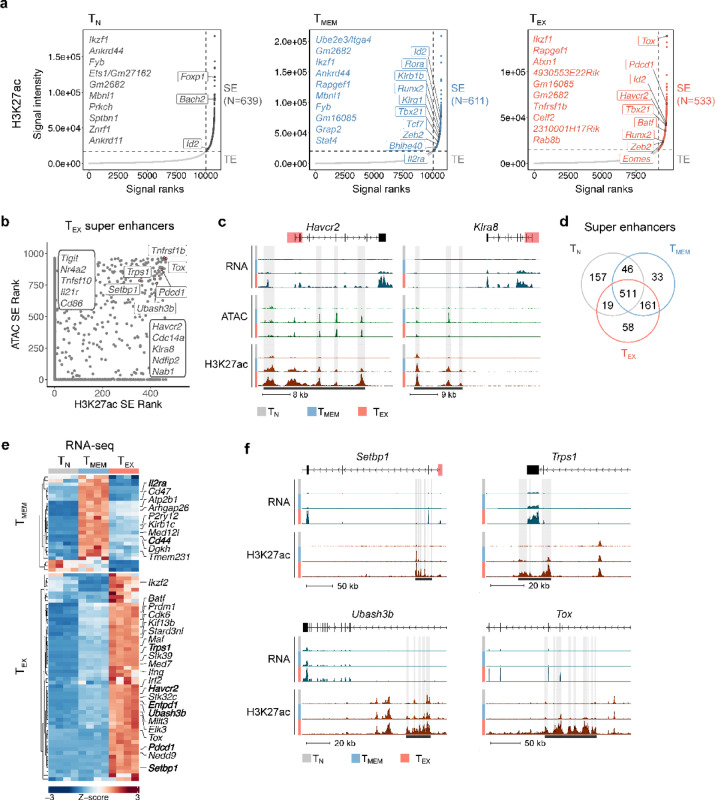
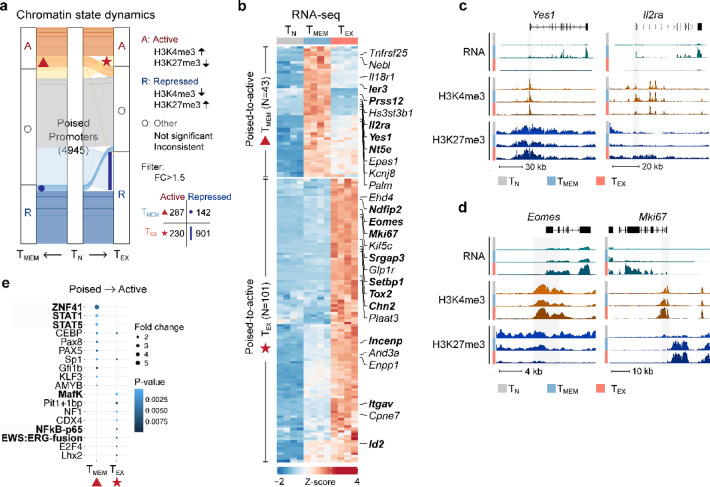
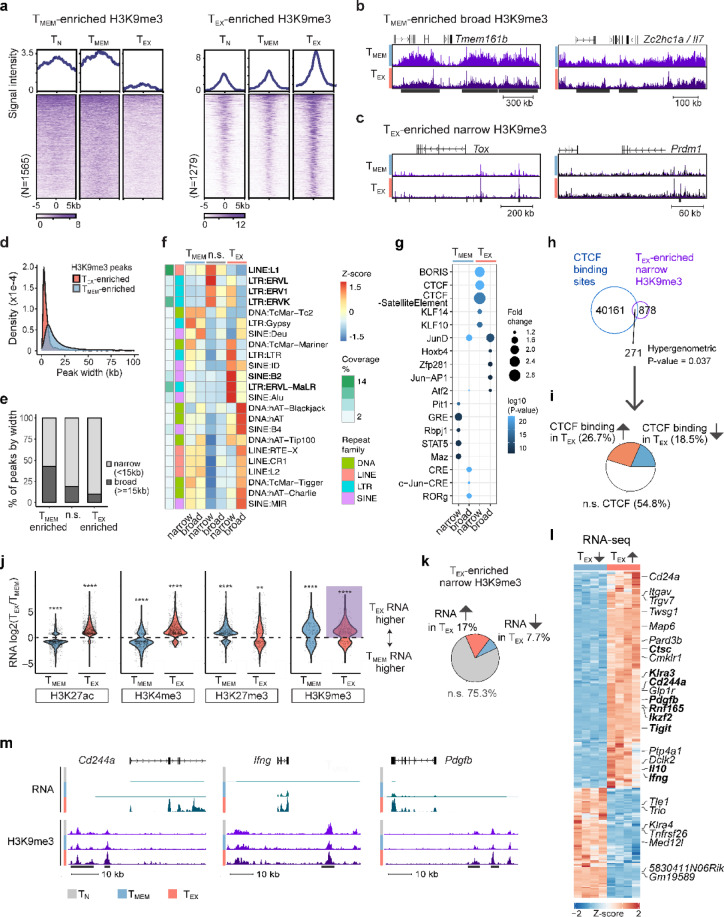
Exhausted CD8 T cells (TEX) arising during chronic infections and cancer have reduced functional capacity and limited fate flexibility that prevents optimal disease control and response to immunotherapies. Compared to memory (TMEM) cells, TEX have a unique open chromatin landscape underlying a distinct gene expression program. How TEX transcriptional and epigenetic landscapes are regulated through histone post-translational modifications (hPTMs) remains unclear. Here, we profiled key activating (H3K27ac and H3K4me3) and repressive (H3K27me3 and H3K9me3) histone modifications in naive CD8 T cells (TN), TMEM and TEX. We identified H3K27ac-associated super-enhancers that distinguish TN, TMEM and TEX, along with key transcription factor networks predicted to regulate these different transcriptional landscapes. Promoters of some key genes were poised in TN, but activated in TMEM or TEX whereas other genes poised in TN were repressed in TMEM or TEX, indicating that both repression and activation of poised genes may enforce these distinct cell states. Moreover, narrow peaks of repressive H3K9me3 were associated with increased gene expression in TEX, suggesting an atypical role for this modification. These data indicate that beyond chromatin accessibility, hPTMs differentially regulate specific gene expression programs of TEX compared to TMEM through both activating and repressive pathways.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: E.J.W. is a member of the Parker Institute for Cancer Immunotherapy. E.J.W. is an advisor for Arsenal Biosciences, Coherus, Danger Bio, IpiNovyx, New Limit, Marengo, Pluto Immunotherapeutics, Prox Bio, Related Sciences, Santa Ana Bio, and Synthekine. E.J.W. is a founder of Arsenal Biosciences, Danger Bio, Prox Bio and holds stock in Coherus. J.R.G. is a consultant for Arsenal Biosciences, Cellanome, GVM1, and Seismic Therapeutics. The remaining authors declare no competing interests. Ethical approval: This study is reported in accordance with ARRIVE guidelines.
Figures
Similar articles
-
Bath: a Bayesian approach to analyze epigenetic transitions reveals a dual role of H3K27me3 in chondrogenesis.Epigenetics Chromatin. 2025 Jun 27;18(1):38. doi: 10.1186/s13072-025-00594-6. Epigenetics Chromatin. 2025. PMID: 40571950 Free PMC article.
-
Epigenomic preconditioning of peripheral monocytes determines their transcriptional response to the tumor microenvironment.Genome Med. 2025 Jul 23;17(1):82. doi: 10.1186/s13073-025-01511-y. Genome Med. 2025. PMID: 40702585 Free PMC article.
-
Dynamic epigenomic landscape and gene regulatory networks during embryonic development in Pacific white shrimp (Litopenaeus vannamei) as revealed by histone modification profiling using CUT&Tag.Epigenetics Chromatin. 2025 Aug 4;18(1):50. doi: 10.1186/s13072-025-00615-4. Epigenetics Chromatin. 2025. PMID: 40760451 Free PMC article.
-
Bivalent Histone Modifications and Development.Curr Stem Cell Res Ther. 2018;13(2):83-90. doi: 10.2174/1574888X12666170123144743. Curr Stem Cell Res Ther. 2018. PMID: 28117006
-
A narrative review of epigenetic marker in H3K27ac and its emerging potential as a therapeutic target in cancer.Epigenomics. 2025 Mar;17(4):263-279. doi: 10.1080/17501911.2025.2460900. Epub 2025 Feb 21. Epigenomics. 2025. PMID: 39981972 Review.
References
-
- McLane, L. M., Abdel-Hakeem, M. S. & Wherry, E. J. CD8 T cell exhaustion during chronic viral infection and Cancer. Annu. Rev. Immunol.37, 457–495 (2019). - PubMed
-
- Gehart, H. & Clevers, H. Tales from the Crypt: New insights into intestinal stem cells. Nat. Rev. Gastroenterol. Hepatol.16, 19–34 (2019). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
