

Graphene oxide enhances aphid resistance in sorghum via the miR319-SbTCP7-SbLOX3 Pathway
- PMID: 40397692
- PMCID: PMC12310823
- DOI: 10.1111/pbi.70132
Graphene oxide enhances aphid resistance in sorghum via the miR319-SbTCP7-SbLOX3 Pathway
Abstract
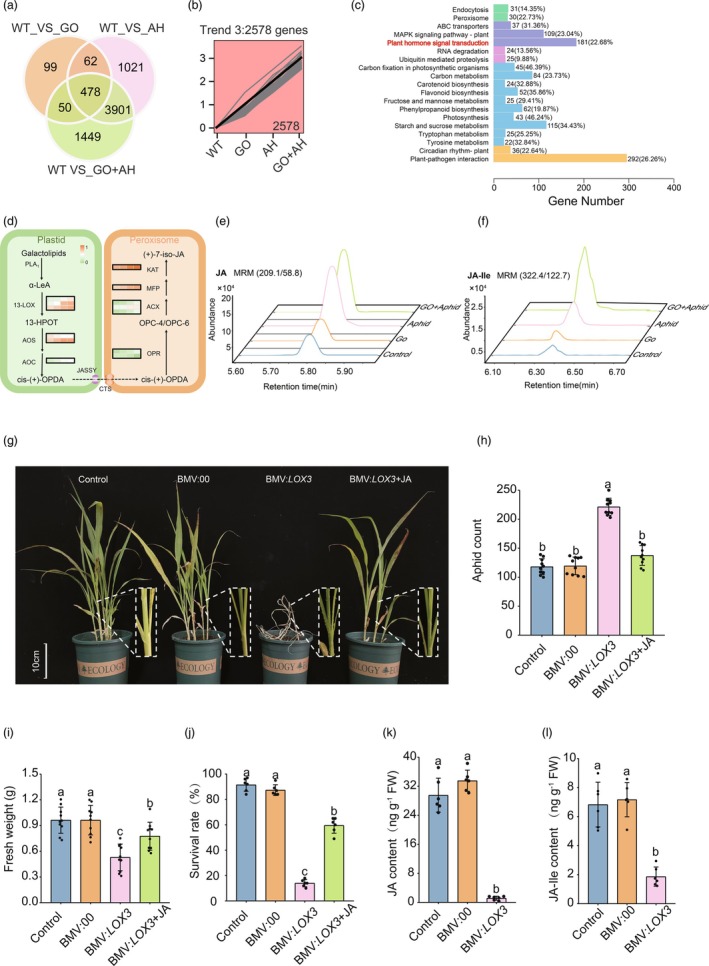
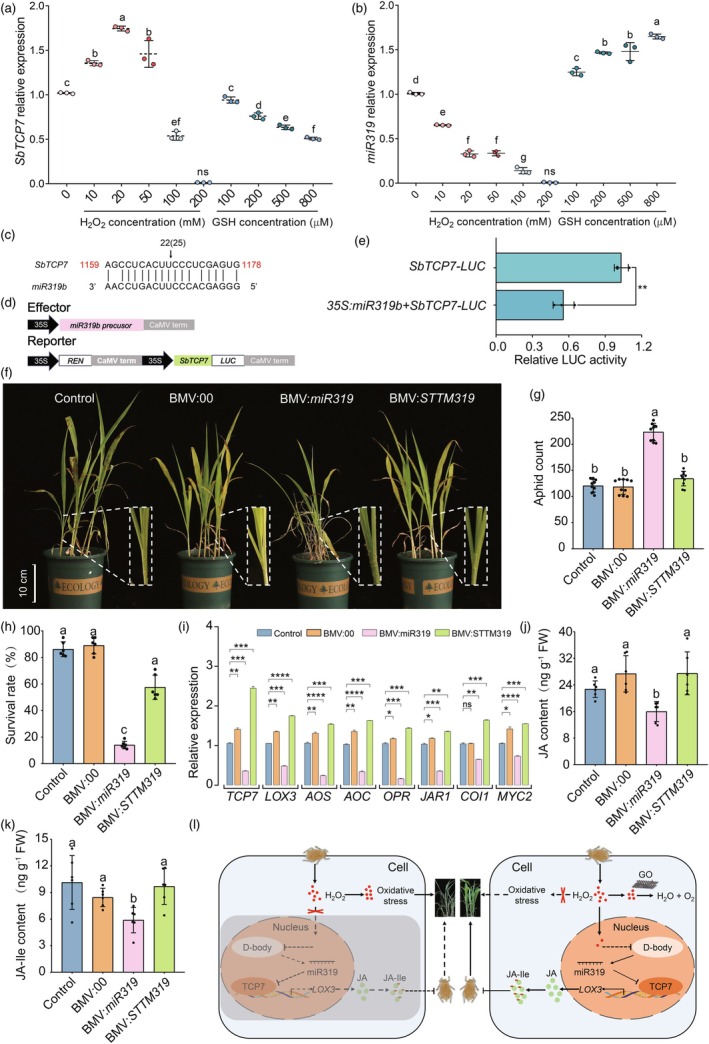
The aphid (Melanaphis sacchari) has emerged as a formidable pest, devastating sorghum plants and highlighting the need for sustainable management strategies. Graphene oxide (GO), as a novel material, has garnered attention for its use in crop cultivation and management, but its effects on biotic stresses remain elusive. Here, we used 10 mg/L GO to spray aphid-stressed sorghum seedlings four times in total. GO exposure reduced 50% H2O2 from the reactive oxygen species (ROS) burst induced by the aphid. Further analysis revealed that GO within the cells acts as a nanozyme, mimicking and enhancing the catalytic activity of the ROS-scavenging system to maintain ROS homeostasis, protecting normal plant growth and development under aphid stress. Moreover, the moderate increase in H2O2 in GO-treated, aphid-infected seedlings blocked the biogenesis of miR319, leading to the induction of its target gene SbTCP7, which in turn activated the transcription of SbLOX3, a rate-limiting enzyme in jasmonic acid (JA) biosynthesis. Subsequent molecular and genetic assays confirmed that the miR319-SbTCP7 module enhances JA metabolism, promoting the accumulation of JA and its active derivative jasmonic acid-isoleucine (JA-Ile) to combat aphids. Our results suggest that GO, as a potential nanozyme, enhances the aphid resistance of sorghum through the miR319-SbTCP7 module to regulate JA synthesis, indicating a novel cultivation strategy for improving pest management via nanomaterials. This frontier research has opened new avenues for crop protection against invasive pests like aphids.
Keywords: aphid; graphene oxide; jasmonic acid (JA); miR319; sorghum.
© 2025 The Author(s). Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Allen, M.J. , Tung, V.C. and Kaner, R.B. (2010) Honeycomb carbon: a review of graphene. Chem. Rev. 110, 132–145. - PubMed
-
- Armstrong, J.S. , Rooney, W.L. , Peterson, G.C. , Villenueva, R.T. , Brewer, M.J. and Sekula‐Ortiz, D. (2015) Sugarcane aphid (Hemiptera: Aphididae): Host range and sorghum resistance including cross‐resistance from greenbug sources. J. Econ. Entomol. 108, 576–582. - PubMed
-
- Borgatta, J. , Ma, C. , Hudson‐Smith, N. , Elmer, W. , Plaza Pérez, C.D. , De La Torre‐Roche, R. , Zuverza‐Mena, N. et al. (2018) Copper based nanomaterials suppress root fungal disease in watermelon (Citrullus lanatus): role of particle morphology, composition and dissolution behavior. ACS Sustain. Chem. Eng. 6, 14847–14856.
-
- Brewer, M.J. , Gordy, J.W. , Kerns, D.L. , Woolley, J.B. , Rooney, W.L. and Bowling, R.D. (2017) Sugarcane aphid population growth, plant injury, and natural enemies on selected grain sorghum hybrids in Texas and Louisiana. J. Econ. Entomol. 110, 2109–2118. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
