

Horizontal gene transfer of molecular weapons can reshape bacterial competition
- PMID: 40397871
- PMCID: PMC12094771
- DOI: 10.1371/journal.pbio.3003095
Horizontal gene transfer of molecular weapons can reshape bacterial competition
Abstract
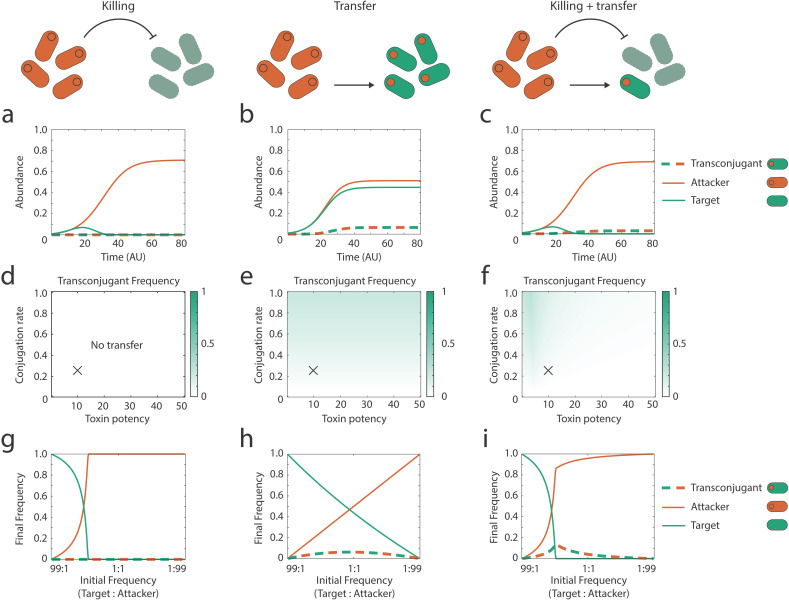
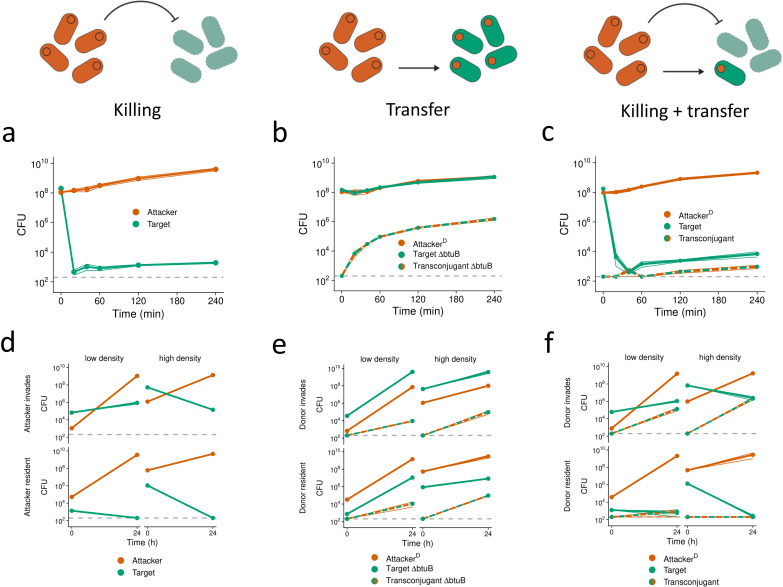
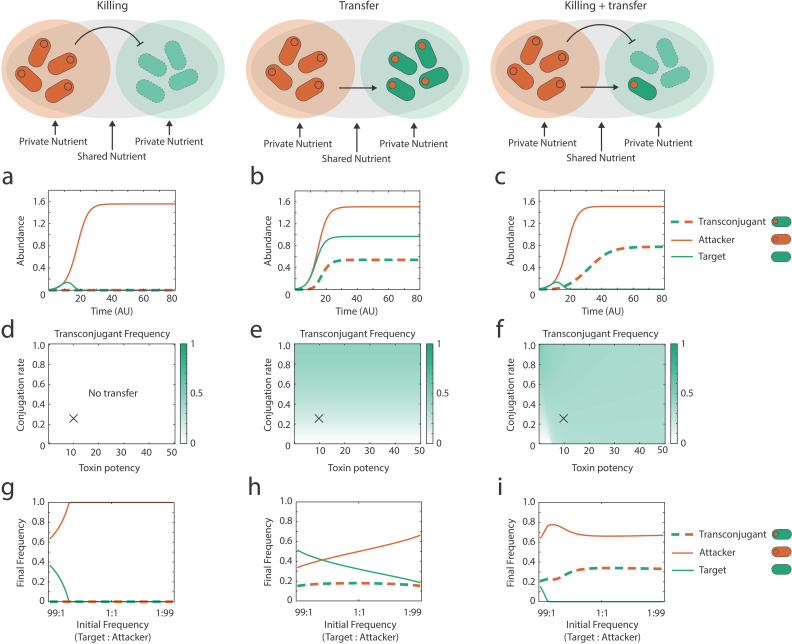
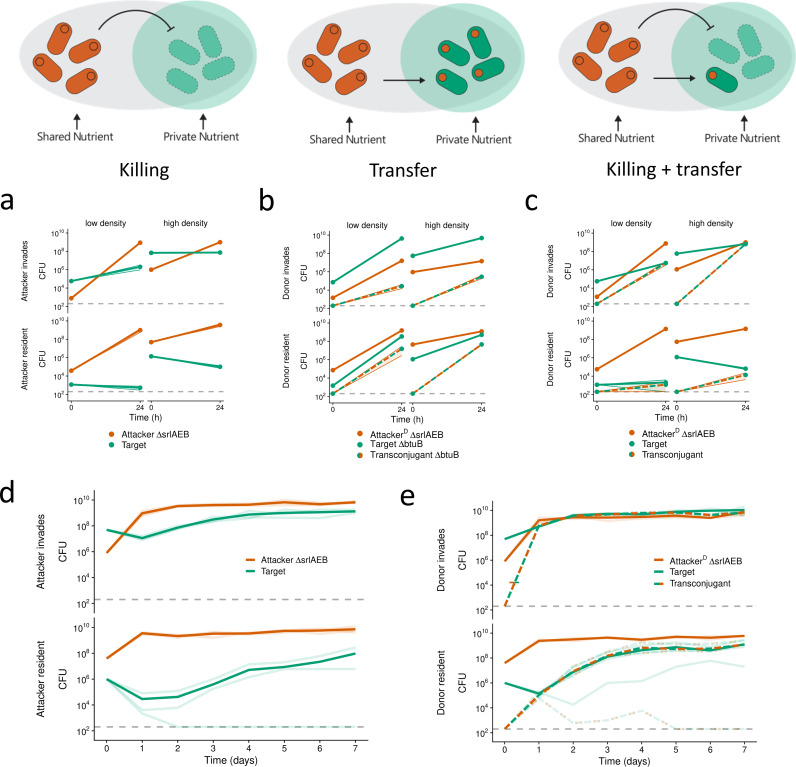
Bacteria commonly use molecular weaponry to kill or inhibit competitors. Genes encoding many weapons and their associated immunity mechanisms can be transmitted horizontally. These transfer events are striking because they appear to undermine bacterial weapons when given to competing strains. Here, we develop an ecological model of bacterial warfare to understand the impacts of horizontal gene transfer. Our model predicts that weapon gene transfer from an attacker to a target strain is possible, but will typically occur at a low rate such that transfer has a negligible impact on competition outcomes. We tested the model empirically using a transmissible plasmid encoding colicin E2, a potent antibacterial toxin produced by Escherichia coli. As predicted by the model, we find that toxin plasmid transfer is feasible during warfare, but the resulting transconjugants remain rare. However, exploring the model further reveals realistic conditions where transfer is predicted to have major impacts. Specifically, the model predicts that whenever competing strains have access to unique nutrients, transconjugants can proliferate and reach high abundances. In support of these predictions, short- and long-term experiments show that transconjugants can thrive when nutrient competition is relaxed. Our work shows how horizontal gene transfer can reshape bacterial warfare in a way that benefits a weapon gene and strains that receive it. Interestingly, we also find that there is little cost to a strain that transfers a weapon gene, which is expected to further enable the horizontal gene transfer of molecular weapons.
Copyright: © 2025 Granato et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Colicin Immunity Proteins of Pathogenic Bacteria Detected by Antibiotic-Induced SOS Response, Plasmid Sequencing, MALDI-TOF-TOF Mass Spectrometry, and Top-Down Proteomic Analysis.Rapid Commun Mass Spectrom. 2025 Mar;39(5):e9964. doi: 10.1002/rcm.9964. Rapid Commun Mass Spectrom. 2025. PMID: 39668116
-
A Novel Mobilizing Tool Based on the Conjugative Transfer System of the IncM Plasmid pCTX-M3.Appl Environ Microbiol. 2020 Aug 18;86(17):e01205-20. doi: 10.1128/AEM.01205-20. Print 2020 Aug 18. Appl Environ Microbiol. 2020. PMID: 32591385 Free PMC article.
-
Escherichia coli K317, formerly used to define colicin group E2, produces colicin E7, is immune to colicin E2, and carries a bacteriophage-restricting conjugative plasmid.J Bacteriol. 1980 Nov;144(2):524-31. doi: 10.1128/jb.144.2.524-531.1980. J Bacteriol. 1980. PMID: 7000748 Free PMC article.
-
Mouse Genetic Background Affects Transfer of an Antibiotic Resistance Plasmid in the Gastrointestinal Tract.mSphere. 2020 Jan 29;5(1):e00847-19. doi: 10.1128/mSphere.00847-19. mSphere. 2020. PMID: 31996415 Free PMC article.
-
Mechanisms of bacterial immunity, protection, and survival during interbacterial warfare.Cell Host Microbe. 2024 Jun 12;32(6):794-803. doi: 10.1016/j.chom.2024.05.006. Cell Host Microbe. 2024. PMID: 38870897 Free PMC article. Review.
Cited by
-
Antibacterial microcins are ubiquitous and functionally diverse across bacterial communities.Nat Commun. 2025 Jul 1;16(1):6048. doi: 10.1038/s41467-025-61151-z. Nat Commun. 2025. PMID: 40595659 Free PMC article.
References
-
- Granato ET, Meiller-Legrand TA, Foster KR. The evolution and ecology of bacterial warfare. Curr Biol. 2019;29(11):R521–37. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
