

Vertex models capturing subcellular scales in epithelial tissues
- PMID: 40397938
- PMCID: PMC12094738
- DOI: 10.1371/journal.pcbi.1012993
Vertex models capturing subcellular scales in epithelial tissues
Abstract
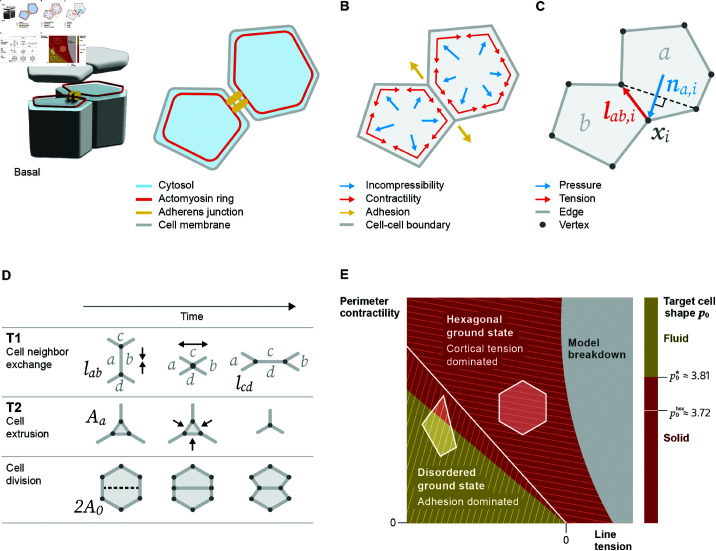
Vertex models provide a robust theoretical framework for studying epithelial tissues as a network of cell boundaries. They have been pivotal in exploring properties such as cell packing geometry and rigidity transitions. Recently, extended vertex models have become instrumental in bridging the subcellular scales to the tissue scale. Here, we review extensions of the model aiming to capture experimentally observed subcellular features of epithelial tissues including heterogeneity in myosin activity across the tissue, non-uniform contractility structures, and mechanosensitive feedback loops. We discuss how these extensions change and challenge current perspectives on observables of macroscopic tissue properties. First, we find that extensions to the vertex model can change model properties significantly, impacting the critical threshold and in some cases even the existence of a rigidity transition. Second, we find that packing disorder can be explained by models employing different subcellular mechanisms, indicating a source of stochasticity and gradual local size changes as common mesoscopic motifs in the mechanics of tissue organization. We address complementary models and statistical inference, putting vertex models in a broader methodological context and we give a brief overview of software packages utilized in increasingly complex vertex model studies. Our review emphasizes the need for more comparative, systematic studies that identify specific classes of vertex models which share a set of well-defined properties, as well as a more in-depth discussion of modeling choices and their biological motivations.
Copyright: © 2025 Lange et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Weaire D, Hutzler S. The physics of foams. Oxford: Oxford University Press; 2000
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
