

GSNOR plays roles in growth, pathogenicity, and stress resistance by modulating mitochondrial protein COX6B S-nitrosylation in Colletotrichum gloeosporioides
- PMID: 40407326
- PMCID: PMC12153361
- DOI: 10.1128/mbio.01269-25
GSNOR plays roles in growth, pathogenicity, and stress resistance by modulating mitochondrial protein COX6B S-nitrosylation in Colletotrichum gloeosporioides
Abstract
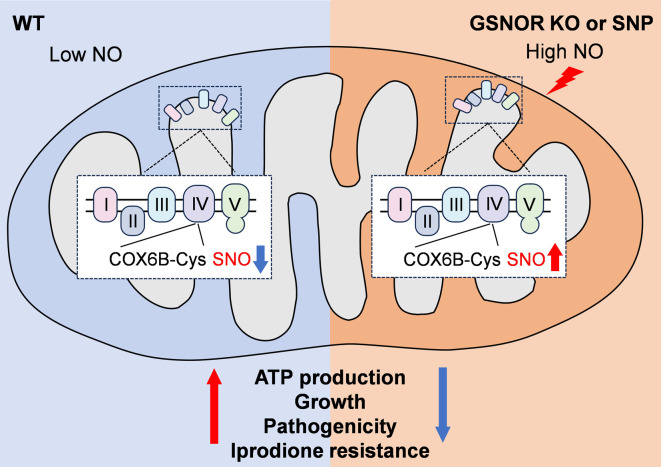
Nitric oxide (NO) serves as a versatile signaling molecule governing diverse biological processes, primarily through post-translational modifications such as S-nitrosylation. The enzyme glutathione S-nitrosoglutathione reductase (GSNOR) plays a central role in NO homeostasis by modulating cellular levels of S-nitrosoglutathione (GSNO), thereby controlling protein S-nitrosylation dynamics. However, the functional significance of GSNOR in fungal pathogenicity remains insufficiently characterized. In this study, we investigated the function of CgGSNOR in the phytopathogenic fungus Colletotrichum gloeosporioides. Deletion of CgGSNOR disrupted nitrosative homeostasis, leading to elevated NO accumulation, increased protein S-nitrosylation levels, and mitochondrial dysfunction as evidenced by reduced ATP production and altered ROS levels. Proteomic and structural analyses identified cytochrome c oxidase subunit 6B (CgCOX6B) as a key target of S-nitrosylation. Functional characterization revealed that CgCOX6B is essential for appressorial turgor maintenance and fungal pathogenicity. Site-directed mutagenesis demonstrated that three conserved cysteine residues (Cys42, Cys62, and Cys73) are critical for CgCOX6B function and are susceptible to S-nitrosylation-induced disruption. Notably, the CgCOX6B knockout strain exhibited increased sensitivity to Iprodione, a widely used fungicide, and this sensitivity was further amplified by NO donor treatment. Together, our findings uncover a GSNOR-dependent redox regulatory axis that links NO signaling, mitochondrial function, and fungal pathogenicity, offering potential targets for antifungal strategies via manipulation of NO signaling networks.IMPORTANCEColletotrichum gloeosporioides is a globally significant fungal pathogen responsible for anthracnose diseases, causing losses across a wide range of crops. Although nitric oxide (NO) signaling and its post-translational regulatory mechanism, S-nitrosylation, are known to play pivotal roles in fungal biology, their specific contributions to pathogenicity remain poorly characterized. This study identifies glutathione S-nitrosoglutathione reductase (GSNOR) as a critical regulator of NO homeostasis in C. gloeosporioides and demonstrates its critical role in regulating fungal growth, conidiation, and pathogenicity. We uncover cytochrome c oxidase subunit 6B (COX6B) as a key target of S-nitrosylation, required for fungal energy metabolism, host infection, and resistance to fungicides. Furthermore, we reveal that exogenous NO supplementation using sodium nitroprusside synergistically enhances the antifungal activity of Iprodione. These findings advance our understanding of redox regulation in fungal pathogenesis and highlight GSNOR and COX6B as promising molecular targets for developing antifungal approaches to reduce crop losses.
Keywords: Colletotrichum gloeosporioides; S-nitrosylation; cytochrome c oxidase subunit 6B (COX6B); fungal pathogenicity; glutathione S-nitrosoglutathione reductase (GSNOR).
Conflict of interest statement
The authors declare no conflict of interest.
Figures









References
-
- Thomas DD, Ridnour LA, Isenberg JS, Flores-Santana W, Switzer CH, Donzelli S, Hussain P, Vecoli C, Paolocci N, Ambs S, Colton CA, Harris CC, Roberts DD, Wink DA. 2008. The chemical biology of nitric oxide: implications in cellular signaling. Free Radic Biol Med 45:18–31. doi: 10.1016/j.freeradbiomed.2008.03.020 - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
