

Bi-allelic variants in POPDC2 cause an autosomal recessive syndrome presenting with cardiac conduction defects and hypertrophic cardiomyopathy
- PMID: 40409267
- PMCID: PMC12256823
- DOI: 10.1016/j.ajhg.2025.04.016
Bi-allelic variants in POPDC2 cause an autosomal recessive syndrome presenting with cardiac conduction defects and hypertrophic cardiomyopathy
Abstract
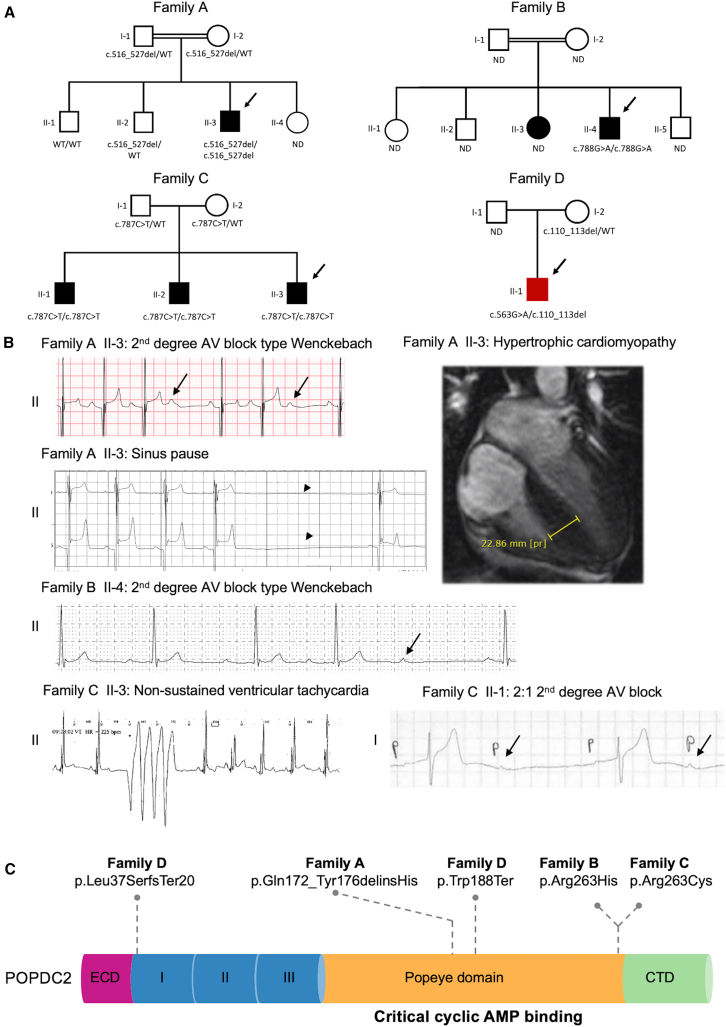
POPDC2 encodes the Popeye domain-containing protein 2, which has an important role in cardiac pacemaking and conduction, due in part to its cyclic AMP (cAMP)-dependent binding and regulation of TREK-1 potassium channels. Loss of Popdc2 in mice results in sinus pauses and bradycardia, and morpholino-mediated knockdown of popdc2 in zebrafish results in atrioventricular (AV) block. We identified bi-allelic variants in POPDC2 in four families with a phenotypic spectrum consisting of sinus node dysfunction, AV conduction defects, and hypertrophic cardiomyopathy. Using homology modeling, we show that the identified variants are predicted to diminish the ability of POPDC2 to bind cAMP. In in vitro electrophysiological studies, we demonstrated that, in contrast with wild-type POPDC2, variants found in affected individuals failed to increase TREK-1 current density. While muscle biopsy of an affected individual did not show clear myopathic disease, it showed significantly reduced abundance of both POPDC1 and POPDC2, suggesting that stability and/or membrane trafficking of the POPDC1-POPDC2 complex is impaired by pathogenic variants in either protein. Single-cell RNA sequencing from human hearts demonstrated that co-expression of POPDC1 and POPDC2 was most prevalent in AV node, AV node pacemaker, and AV bundle cells. Using population-level genetic data of more than 1 million individuals, we show that none of the familial variants were associated with clinical outcomes in heterozygous state, suggesting that heterozygous family members are unlikely to develop clinical manifestations and therefore might not necessitate clinical follow-up. Our findings provide evidence for bi-allelic variants in POPDC2 causing a Mendelian autosomal recessive cardiac syndrome.
Keywords: AV conduction defects; cardiac arrhythmia; hypertrophic cardiomyopathy; population genetics; sinus node disease.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L. Monserrat is a shareholder in Dilemma Solutions SL. D.A.C. is an employee of and may own stock in GeneDx. H.M.A., V.T., G.S., E.V.I., H.H., D.F.G., A.T.S., and K.S. report employment at deCODE Genetics during the conduct of the study. C.E. reports grants from Abbott Diagnostics and Novo Nordisk outside the submitted work. K.U.K. reports research support from Intermountain Foundation during the conduct of the study. L.N. reports a stock option grant from Culmination Bio. H.B. reports lecture fees from Amgen, MSD, Sanofi Avensis, Bristol Myers Squibb, and Pfizer; grants from Novo Nordic Foundation; and another from Novo Nordic Foundation (shares) outside the submitted work.
Figures





Update of
-
Biallelic variants in POPDC2 cause a novel autosomal recessive syndrome presenting with cardiac conduction defects and variable hypertrophic cardiomyopathy.medRxiv [Preprint]. 2024 Jul 5:2024.07.04.24309755. doi: 10.1101/2024.07.04.24309755. medRxiv. 2024. Update in: Am J Hum Genet. 2025 Jul 3;112(7):1681-1698. doi: 10.1016/j.ajhg.2025.04.016. PMID: 39006410 Free PMC article. Updated. Preprint.
References
-
- Wilde A.A.M., Semsarian C., Márquez M.F., Sepehri Shamloo A., Ackerman M.J., Ashley E.A., Sternick E.B., Barajas-Martinez H., Behr E.R., Bezzina C.R., et al. European Heart Rhythm Association (EHRA)/Heart Rhythm Society (HRS)/Asia Pacific Heart Rhythm Society (APHRS)/Latin American Heart Rhythm Society (LAHRS) Expert Consensus Statement on the state of genetic testing for cardiac diseases. Europace. 2022;24 doi: 10.1093/europace/euac030. 1307–136. - DOI - PubMed
-
- Schindler R.F.R., Scotton C., Zhang J., Passarelli C., Ortiz-Bonnin B., Simrick S., Schwerte T., Poon K.L., Fang M., Rinné S., et al. POPDC1S201F causes muscular dystrophy and arrhythmia by affecting protein trafficking. J. Clin. Investig. 2016;126:239–253. doi: 10.1172/JCI79562. - DOI - PMC - PubMed
-
- Froese A., Breher S.S., Waldeyer C., Schindler R.F.R., Nikolaev V.O., Rinné S., Wischmeyer E., Schlueter J., Becher J., Simrick S., et al. Popeye domain containing proteins are essential for stress-mediated modulation of cardiac pacemaking in mice. J. Clin. Investig. 2012;122:1119–1130. doi: 10.1172/JCI59410. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
